


- Accueil
- Volume 15 (2011)
- numéro 3
- Biosensors in forensic sciences


Visualisation(s): 12105 (410 ULiège)
Téléchargement(s): 281 (4 ULiège)

Biosensors in forensic sciences

Notes de la rédaction
Received on July 8, 2010; accepted on February 8, 2011
Résumé
Les biodétecteurs en sciences forensiques. Un biodétecteur est un dispositif qui emploie des organismes biologiques pour surveiller la présence de divers produits chimiques dans un secteur particulier. La méthode traditionnelle de détection des substances volatiles, qui est employée par la police, se base sur les capacités olfactives des chiens. Ce concept d'employer des chiens pour détecter certaines substances n'est pas récent. Cependant, l'utilisation de ces chiens présente certaines limites telles que le cout de formation, une longue période d'apprentissage, etc. Ainsi, les chercheurs se sont penchés sur l'utilisation d'autres biodétecteurs tels que des rats, les dauphins, les abeilles, les parasitoïdes dans la détection d'explosifs, de drogues ou de cadavres. Contrairement aux mammifères, les insectes présentent plusieurs avantages. Ceux-ci sont très sensibles, sont peu couteux et peuvent être conditionnés avec une vitesse impressionnante pour une tâche spécifique de produit-détection. D'ailleurs, l'utilisation des insectes comme biodétecteurs pourrait être préférée dans les scénarios qui sont considérés comme trop dangereux pour des mammifères. Le but de cette synthèse bibliographique est de fournir l'ensemble des biodétecteurs utilisés en sciences forensiques.
Abstract
A biosensor is a device that uses biological materials to detect and monitor the presence of specific chemicals in an area. Traditional methods of volatile detection used by law enforcement agencies and rescue teams typically consist of reliance on canine olfaction. This concept of using dogs to detect specific substances is quite old. However, dogs have some limitations such as cost of training and time of conditioning. Thus, the possibility of using other organisms as biosensors including rats, dolphins, honeybees, and parasitic wasps for detecting explosives, narcotics and cadavers has been developed. Insects have several advantages unshared by mammals. Insects are sensitive, cheap to produce and can be conditioned with impressive speed for a specific chemical-detection task. Moreover, insects might be a preferred sensing method in scenarios that are deemed too dangerous to use mammals. The purpose of this review is to provide an overview of the biosensors used in forensic sciences.
Table des matières
1. Introduction
1Biological sensors, biosensors or biodetectors are defined as analytical devices incorporating a biological material (e.g., tissue, microorganisms, insects, animals, enzymes, antibodies, plants), a biologically derived material (e.g., recombinant antibodies, engineered proteins) or a biomimic (e.g., synthetic receptors, combinatorial ligands) intimately associated with or integrated within a physicochemical detector or transducing microsystem, which may be optical, electrochemical, thermometric, piezoelectric, magnetic or micromechanical (Weetall, 1996). Biosensors usually yield an electronic signal which is proportional to the concentration of a specific or group of analytes. Biosensors have been applied to a wide variety of analytical problems including medicine (Clark et al., 1962), biomedical research (Lanzoni et al., 2009), drug discovery (Harper et al., 2007), environment (Amine et al., 2006; Badihi-Mossberg et al., 2007), food (Amine et al., 2006), process industries (Tokarskyy et al., 2008), security (Harper et al., 2007), and defense (Harper et al., 2007). Most of these biological sensors are used for their intrinsic properties, but scientists produce also genetically modified organisms as biodetectors (Weetall, 1996). Because of their exceptional capabilities, including high specificity and sensitivity, rapid response, low cost, relatively compact size and user-friendly operation, biosensors have become an important tool for detection of chemical and biological components for clinical, food, law and environmental monitoring (Weetall, 1996).
2Forensic sciences are the application of a broad spectrum of sciences to answer questions of interest to a legal system (Ricciuti, 2007). This may be in relation to a crime or a civil action. Forensic sciences include several under divisions like forensic entomology, forensic toxicology, forensic anthropology, DNA analysis, criminalistics, and aim to analyze criminal evidences (Ricciuti, 2007). The results are further presented as accurately and precisely as possible in a court of law (Ricciuti, 2007). Forensic sciences do not use only chemical technologies to solve a crime; they use also vertebrates or invertebrates as chemical detectors. This use is based on the olfactory capacities of vertebrates and invertebrates to detect volatiles from a human, narcotics or explosives.
3In the field of forensic sciences, law enforcement agencies and rescue teams worldwide use biosensors including dogs and honeybees to locate corpses, drugs or explosives.
4In this review, we specifically provide an overview of the biosensor systems used in forensic sciences.
2. Principle of the biosensors used in forensic sciences
2.1. Associative learning
5Associative learning is common throughout the animal kingdom, from invertebrates to vertebrates (Tomberlin et al., 2005). Learning ability has evolved to adjust the behavioral response of animals to changing ecological conditions (Stephens, 1993). To condition animals, two techniques of learning are used, i.e. they must go through a phase of training.
6Operant conditioning, or instrumental learning, is a form of associative learning that occurs through rewards and punishments for behavior (Skipper, 1938). Through operant conditioning, an association is made between a behavior and a consequence for that behavior. There are two types of operant conditioning:
7- Positive Reinforcement: a particular behavior is strengthened by the consequence of experiencing a positive condition (Bernstein et al., 2008). For example, a hungry rat presses a bar in its cage and receives food. The food is a positive condition for the hungry rat. The rat presses the bar again, and again receives food. The rat's behavior of pressing the bar is strengthened by the consequence of receiving food.
8- Negative Reinforcement: a particular behavior is strengthened by the consequence of stopping or avoiding a negative condition (Bernstein et al., 2008). For example, a rat is placed in a cage and immediately receives a mild electrical shock on its feet. The shock is a negative condition for the rat. The rat presses a bar and the shock stops. The rat receives another shock, presses the bar again, and again the shock stops. The rat's behavior of pressing the bar is strengthened by the consequence of stopping the shock.
9Both positive and negative reinforcement strengthen behavior. In other words, consequences – whether good or bad – determine if a behavior is maintained or not. For example, dolphins get a fish for doing a trick. Because the animal wants to gain that good thing again, it will repeat the behavior that seems to cause that consequence (Bernstein et al., 2008).
10The second technique of learning which is used in forensic sciences is the classical conditioning, which is synonymous with Pavlovian or respondent conditioning. It is a form of associative learning that was first demonstrated by Ivan Pavlov (Pavlov et al., 1960). The typical procedure for inducing classical conditioning involves presentations of a neutral stimulus along with a stimulus of some significance (Figure 1). The neutral stimulus could be any event that does not result in an overt behavioral response from the organism under investigation. This is referred as a conditioned stimulus. Conversely, presentation of the significant stimulus necessarily evokes an innate response. This response is called the unconditioned stimulus. If the conditioned stimulus and the unconditioned stimulus are repeatedly paired, eventually the two stimuli become associated and the organism begins to produce a behavioral response to the conditioned stimulus (Pavlov et al., 1960). This mechanism called conditioned response in which the response to the conditioned stimulus is greater than the original baseline response elicited by the conditioned stimulus prior to conditioning. This type of learning has the most potential for application toward the development of biological sensors (Tomberlin et al., 2008).
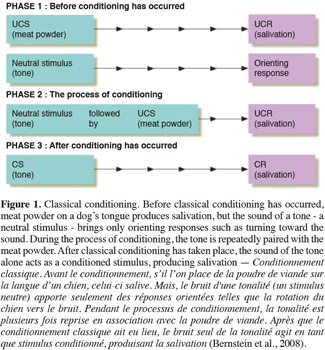
11These two conditionings are based on the olfactory system of animals.
2.2. Olfaction
12Chemical communication or chemoreception is the principal means of communication for many groups of animals (Banaigs, 2002). Chemoreception is the process of detecting chemical compounds by a living organism and involves molecular interactions with olfactory neurons by molecules that have moderate molecular weight, low polarity, particular water solubility, high vapor pressure, and lipophilicity (Meierhenrich et al., 2004). Olfaction allows the animals to detect and recognize the chemical signals of their environment which enable them to communicate with their congenerics, to locate a source of food, to seek a site of laying, to establish inter-individual, social and sexual relations (Banaigs, 2002). The detection of an odor by the olfactory organs causes, if it is identified and recognized, the release or the modification of the behavior (Banaigs, 2002). This olfactory capacity will be used in the learning mechanism to create biosensors.
13The olfactory mechanism is organized in three stages: reception, transduction and integration of the sensory signal (Brossut, 1996).
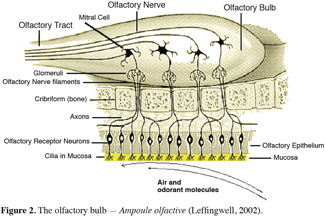
14In vertebrates, odorants generally first enter the olfactory system by passing through the external nares of vertebrates where warming and humidification occur before passing through a set of cartilaginous flap called turbinates that act to increase the surface area of the epithelium (Leffingwell, 2002; Meierhenrich et al., 2004). Then, odorants are thought to be associated with odorant-transport proteins that help transport the odorant from the inhaled air stream through the mucus to the cilia of the olfactory neurons (Figure 2). The odorant binds with a receptor protein (embedded in the phospholipid surface membrane) which activates this one and a multitude of molecular interactions. On this level, the transduction of the chemical signal in an electrical signal is produced (Stengl et al., 1999). Then, the electrical signal is sent via the axon, through the cribiform plate to the glomerulus and mitral cells, then on through the olfactory tract and into the brain where the odor is interpreted (Leffingwell, 2002; Meierhenrich et al., 2004).
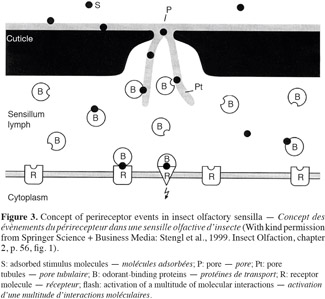
15In insects, when a stimulus molecule comes into contact with the olfactory sensilla localized on the antenna, legs and maxillary palps of the insect, it will be adsorbed and the molecule diffuses from the hair surface through the cuticular pores and pore tubules to the hair lumen (Figure 3) (Keil, 1999). There, the odorants are taken up by the odorant-binding proteins (OBP) and are transported through the aqueous sensillum lymph until they reach a receptor molecule of the outer dendritic membrane. Odorant fixation on the receptor molecule activates results in activation and a multitude of molecular interactions. On this level, the transduction of the chemical signal in an electrical signal is produced (Stengl et al., 1999). Then, the electrical signal is sent via the axon until the primary education processing center of the central nervous system: the antennal lobe. From there, information are transmitted to the brain (protocerebrum) where they will be integrated and memorized to produce a behavioral reaction (Brossut, 1996).
3. Biosensors
3.1. Vertebrate
16Dog. The most widely deployed detector to date is Canis familiaris Linnaeus, better known as the common dog (Harper et al., 2007). Sniffing dogs are used ubiquitously by law enforcement and private agencies for detection of many different items. The most common items of forensic interest for which canines have been deployed to locate are drugs, ignitable liquid residues, explosives, human remains, and human scent (disaster survivors) (Harper et al., 2007; Oesterhelweg et al., 2008). In addition, trained dogs have the ability to detect decomposing bodies beneath running water (Cornaz, 2008). The use of the canine as a detector is based on the well-established reliability and impressive selectivity and sensitivity associated with dog's sense of smell (Harper et al., 2007). No one really knows how sensitive a dog's nose is, but scientists note that a dog's sensitivity to odor is of 100 times at 1,000 times greater than that of a human (Seuter, 2003). Moreover, they can track directions, have long-term olfactory memory and portray extreme discriminative ability (Harper et al., 2007).
17The training of the dogs begins with the basic conditioning (Cornaz, 2008). During two years, they will be familiarized with the search of objects and people, especially with obedience. Then, the various junctions intervene: drugs, explosives, corpses or missing people. These two last disciplines take the most time to train dogs (Cornaz, 2008). The training of sniffing dogs is based on the recognition of a particular odor and on the learning to alter their behaviors when certain odors denoting specific substances are found. The training is based on positive reinforcement which consists of food. There exist several methods but the details of those are not revealed by the police. During the training, when the dog detects the targeted object with the odor of corpse, drugs or explosives, the trainer gives a reward (food) to the dog to render comprehensible to him its satisfaction. Thus, the dog will associate the odor of the object with reward. The training progresses while burying corpses, drugs or explosives and by modifying the search areas with different environmental conditions. To control the correct memorizing of the odors, the dog is trained daily. The training program is only a guide and each breed and age of dog and condition need to be considered (Rebmann et al., 2000).
18Cues used by a dog to indicate forensic materiel depend on the trainer. There are many different ways a dog can be trained to communicate an alert to its trainer (Lasseter et al., 2003). The dog can give an aggressive or passive alert. An aggressive alert is one in which the dog digs at the site of the scent of interest as human remains, drugs, and/or explosives. A passive alert is when the dog lays down on the site of interest or jumps on the trainer to indicate the presence of remains, drugs or explosives. It is better to give a passive alert than an aggressive alert because crime scenes can be disturbed or altered by the anxious digging of a dog. Specifically, cadaver dogs are trained to find scents, not bodies (Lasseter et al., 2003).
19Canines that detect human remains, commonly referred to as cadaver dogs, are trained in detecting the odor of decomposing bodies. They are used in a variety of forensic contexts, including search and discovery of human cadavers, body parts, or body fluids (Rebmann et al, 2000). The scent picture changes as the body progresses through the decomposition stages. It is important to train dogs on the all stages of decomposition.
20Rescue dogs are trained to detect odors associated with living people such as urine, evaporated perspiration, respiratory gases, or decomposition gases released by bacterial action on human skin or tissues (SARDUS, 1992). Rescue dogs can be classified according to whether they "scent discriminate", and under what conditions they can work. Scent discriminating dogs have proven their ability to alert only on the scent of an individual person, after being given a sample of that person's scent. Non-scent discriminating dogs alert on or follow any scent of a given type, such as any human scent or any cadaver scent (Rebmann et al., 2000). Rescue dogs can be trained specifically for rubble searches, for water searches, and for avalanche searches (SARDUS, 1992).
21Narcotic detection canines are expected to face a predictable lineup of five or six drug odors, the explosive detection canine is expected to face dozens of different potential explosive products during its service (Given, 2003). Narcotics detection canines are typically trained on cocaine, heroin and marijuana. They may be trained on additional drugs, including methamphetamine, ecstasy, hashish, opium, mescaline, and LSD (N, N-diéthyllysergamide), depending on the training agency and the locations where they are deployed (Given, 2003). Explosive detection canines are currently trained on a wide variety of samples ranging from half a dozen samples to upward of 20 (Harper et al., 2007). Using at least one representative sample from each explosive chemical class would require an acid salt such as ammonium nitrate, an aromatic nitro such as TNT (trinitrotoluene), a nitrate ester such as PETN (1,3-dinitrooxy-2,2-bis(nitrooxymethyl) propane), a nitramine such as RDX (cyclonite), an aliphatic nitro such as DMNB (2,3-Dimethyl-2,3-dinitrobutane), a peroxide such as TATP (triacetone triperoxide), and representative black and smokeless powders (Yinon et al., 1996; Harper et al., 2007). All these sniffer dogs should not be used to do other types of work they were not trained for.
22The major disadvantages of detector dogs are their limited duty cycles (necessitating multiple teams for continuous coverage) and possibility of operator influence (because the trainer is dealing with an intelligent animal). The canine's ability to work is also affected by temperature, humidity and wind (Seuter, 2003). The worst conditions for using dogs are when it is hot and dry with little or no air movement and when it is raining or snowing heavily (Lasseter et al., 2003). The most difficult and time-consuming step in the development of a sniffing dog was limiting the negative influence of environmental distractions (audio, visual, olfactory), which disrupted its concentration and could evoke undesired behavior (Yinon et al., 1996). Canine/trainer teams require a great deal of commitment, and dog must also be accompanied by specific trainer in order to work properly (Otto et al., 2002). In addition, dogs are instructed to give the same alert for each explosive or drug; trainers are unaware of the exact type of explosives, drugs detected. Also, dogs need to be in close proximity to sources in order to sample scent; therefore it may be necessary to deploy multiple teams for larger areas as it may take awhile to sweep (Settles et al., 2001). However, there is a possibility that teams may interfere with one another. For smaller areas, accessibility is another issue of which security managers must be aware (Seuter, 2003). If dogs are unable to reach into tight spaces, there is a danger of missing potential hiding spots. This is perhaps one of the most serious limitations of canine detectors. Another disadvantage is the cost of training, the cost for the maintenance of the welfare of the dogs, a hope of use not exceeding the 6-8 years (Harper et al., 2007).
23Rat/Rodents. Given the limitations of canines, researchers are studying the use of other animals in detection of explosives and drugs, even in the presence of distracting odors such as engine oil and almond extract (Otto et al., 2002). The idea of using rats to detect substances has been studied previously by few researchers. Nolan et al. (1978) and Weinstein et al. (1992) studied devices that rewarded rats for proper explosive odor detection using electrical brain stimulation. For this, they used operant conditioning. Odors were delivered to rats in conditioning chambers and odor classification was accomplished via typical lever pressing and via direct monitoring of the rat's cortical frequency spectra. Otto et al. (2002) train the rats, so that their location and alerting behaviors can be monitored from afar by computer and/or humans. Under this concept, the trained alerting behavior of rats is remotely monitored by humans and/or computers to determine when the animals detect the scent of interest as cocaine during their search behavior. When the rats find what they are looking for, they raise the alarm by standing on their hind legs, which is detected by computer (Otto et al., 2002). Since 2003, a social enterprise, APOPO, develops a training method to detect mines (TNT) with the African giant pouched rat or Cricetomys gambianus Waterhouse. The procedure consists of a combination of click training and food rewarding. At first, the animals are taught to associate the click sound with a food reward. Then they have to perform certain tasks to get this food reward. After odor imprint, the complexity of their tasks to be performed is gradually increased. Intellectually, the rats are “smart” enough to learn the desired tasks relatively quickly, while being “uncomplicated” enough for learning to be standardized. Food provides a strong and controllable source of motivation and an effective drive for performance. The ability of a rat to detect different concentration of TNT in air was investigated (Yinon et al., 1996). A rat was trained to press a bar when air containing TNT vapor was delivered, and to refrain from pressing a bar when air free of TNT was delivered; 100% detection was obtained for a concentration of 2.44 ng.l-1 TNT vapor in air, 95% detection for a concentration of 1.32 ng.l-1 and 60% for a concentration of 1.07 ng.l-1. At all three concentrations there were no false alarms (Yinon et al., 1996).
24Contrary to dogs, rats do not form social relationships and obedience with people, so are not dependent on a specific trainer (APOPO, 2003). They can get into tighter spaces, too, so they might be better able to find casualties in collapsed buildings. Because rats are small, they can squeeze into small areas and may be able to infiltrate areas that dogs cannot. They are inexpensive, and are relatively easy to procure and maintain. Rats may also be better than dogs at maintaining performance during long periods of repetitive work. In addition, their light weight makes it less likely to set off explosives (APOPO, 2003).
25While claims that the rats can match the sensitivity of dogs appear to be well founded, there are still significant difficulties in deploying rats directly in the field. Rats trained on a variety of chemicals can be useful in detecting different types of explosives; however, other than the fact something dangerous is present, rats are not able to tell the trainer whether smokeless powder or cyclonite was detected (Yinon et al., 1996).
26Other mammalian species. Theoretically, almost any mammal has the potential to be used as a biological detector with the primary limitations associated with the size and the mobility of the mammal as well as its trainability. The U.S. Navy's Marine Mammal Program trains dolphins (Tursiops truncatus Montagu) and the sea lion (Zalophus californianus Lesson) to find and mark the location of underwater objects, especially mines and submerged vehicles (SPAWAR, 1992). Dolphins are essential because their biological sonar is unmatched by hardware sonars in detecting objects in the water column and on the sea floor (SPAWAR, 1992). Sea lions are used because they have very sensitive underwater directional hearing and exceptional vision in low light conditions. Both of these species are highly reliable, adaptable, and trainable marine animals. They are trained with operant conditioning, emphasizing the use of positive reinforcement (SPAWAR, 1992).
3.2. Invertebrate
27The use of animals to detect volatiles from drugs, explosives or humans has long been recognized and utilized by law enforcement. However, the ability to learn and detect odors of human, drugs or explosives is not limited to vertebrates. Insects are highly sensitive (parts-per-trillion), flexible, portable and cheap to reproduce, and it is easy to condition them to detect target odorants (Rains et al., 2008). Moreover, insects can be conditioned with impressive speed (few minutes to two days) for a specific chemical-detection task and they do not require trainer in the field (Shaw et al., 2005; Rains et al., 2008). Some of their limitations are that insects do not fly at night, in heavy rain, or in cold weather (Harper et al., 2007).
28Forensic entomology is the broad field where arthropod science and the judicial system interact (Hall et al., 2009). In regards to medico-legal fields, insects have primarily been used to determine the time of death, also known as the post-mortem interval (PMI) (Amendt et al., 2004; Wyss et al., 2006; Gennard, 2007). Insects can also bring information in cases of abuse or neglect of children or elderly (Benecke et al., 2001; Gennard, 2007), providing information on the causes of death or the identity of victims (Benecke et al., 2001; Bourel et al., 2001; Gupta et al., 2004; Gennard, 2007). But, the use of insects like biosensors or biodetectors was not yet considered in forensic entomology. However, one of the big challenges of criminalistic, and more particularly of the law enforcement, is to develop efficient techniques to locate corpses, drugs and explosives. Because of the sensitivity of their olfactory system, it appears that insects also might be used to develop novel methods for detecting and locating chemicals associated with decomposition, drugs and explosives (Rains et al., 2004; Tomberlin et al., 2005).
29Apis mellifera Linnaeus. Learning behavior of honeybees has been studied extensively over the past 40 years because they can learn very rapidly many different cues associated with rewards (Menzel et al., 1996; Scheiner et al., 2001).
30Honeybees learn to associate odors with sucrose reward in a Pavlovian conditioning protocol (Hammer et al., 1995; Menzel et al., 1996; Davis et al., 2005; Carcaud et al., 2009). A hungry bee reflexively extends its proboscis when antennal or proboscis contact chemoreceptors are stimulated by sucrose. The proboscis extension response (PER) can be conditioned with a single conditioning trial by pairing an odor with sucrose applied to antennae and proboscis or with repeated conditioning trials where odor and sucrose are paired once per trial. In PER conditioning, the odor represents the conditioned stimulus and sucrose the unconditioned stimulus (Hammer et al., 1995; Menzel et al., 1996; Carcaud et al., 2009). Bees are conditioned in two hours to seek out buried explosives such as TNT; C4; TATP; 2, 4-DNT; and RDX and drugs materials like methamphetamine and cocaine (Davis et al., 2005; Rains et al., 2008).
31Microplitis croceipes Cresson. Parasitic wasps detect and learn numerous chemical cues associated with their host and food resources, and use them in order to forage more effectively (Tertuliano et al., 2004). Microplitis croceipes (Hymenoptera: Braconidae) a relatively specialized endoparasitoid of three highly polyphagous larval hosts, Helicoverpa zea, Heliothis virescens and Heliothis subflexa (Lepidoptera: Noctuidae) were conditioned with classical Pavlovian to associate several chemicals that they would not encounter in their natural foraging (Olson et al., 2003; Tomberlin et al., 2005; Tomberlin et al., 2008). These compounds were octanal, diisopropyl aminoethanol, two compounds of decaying process: cadaverine and putrescine (Takasu et al., 2007) and three volatile compounds of explosives: cyclohexanone, 2,4- and 3,4-dinitrotoluene (Olson et al., 2003; Tomberlin et al., 2005). This parasitic wasp species learned to associate odors with food resources and subsequently exhibits a characteristic food-seeking behavior when encountering the learned odor. Exposures to food and odor repeated three times with a 10 s training session was sufficient to obtain wasp conditioned close to 80% (Tertuliano et al., 2004; Tomberlin et al., 2005; Tomberlin et al., 2008). When wasps responded to the odor of conditioning, they showed typical food-searching behavior. Wasps rubbed their antenna, lowered the head, and mouthpart extension bringing the labium (Takasu et al., 2007).
32Manduca sexta Linneaus. Detection of explosives is also possible with Manduca sexta Linnaeus moths (Lepidoptera: Sphingidae). M. sexta can be trained to respond with a feeding behavior when exposed to odor signatures from explosives using Pavlovian conditioning (King et al., 2004). Briefly, conditioning is achieved by repeated (usually six) pairings of the target odor followed by food. The adults moths are trained individually, with each trial taking about 15 seconds to perform. This learned response appears to be remembered by the animal for the duration of its adult lifespan (1-2 weeks).
33Detector system. Insects can be used as either free-moving or restrained organisms to detect chemical.
34Free-moving detectors. Free-moving insects are allowed to move towards odor sources without being constrained in a device and are tracked during flight or after arrival to the odor source (Rains et al., 2008). Tracking methods enable the locations of the organisms to be known. Conditioned honeybees are tracked using “light detection and ranging” (LIDAR) measurements (Shaw et al., 2005). LIDAR is a technique that measures the properties of scattered light to determine the distance to the object that scatters the light. Further studies enhanced the LIDAR detection of free-flying honeybees by measuring the back-scattered return signal caused by the wing-beat modulation (Rains et al., 2008).
35Two of the most promising methods to track conditioned insects once they have located the source of the odorant are harmonic radar and radio telemetry. Harmonic radar involves the placement of a transponder on the insects (Riley et al., 2002). The transponder consists of an antenna and electronics that use the incoming radar signal as an energy source. When the transponder detects a radar transmission, it replies by immediately emitting a pulse on another frequency that is a harmonic of the incoming signal. Consequently, it does not require a battery on the insect, thus reducing the weight being carried. However, it is difficult to track insect movement in some environments that can disrupt the transponder signal, such as areas with heavy vegetation. Currently, harmonic radar detection is limited to insect species that are large enough to carry the transponder such as honeybees and moths (Riley et al., 2002). Radio telemetry requires that a radio transmitter is mounted on the insect, which is then tracked with a receiver (Rains et al., 2008).
36The free-moving detectors worked very well in small, outdoor areas, where security guards could easily see where they were swarming, but were harder to track when they were used to detect odors in large, uncontained spaces.
37Restrained organisms. The behavioral response of insects to a conditioned stimulus can also be observed in insects that are restrained within a detection device.
38Inscentinel Ltd (Davis et al., 2005), an enterprise in the UK, uses a detection device in which three honeybees are held in cassettes, with the head of each bee protruding so that the proboscis is easily observed. Air samples are brought into the device and passed over the bee antennae, and a vision system measures the proboscis extension response. This response can be measured visually or electronically.
39Rains et al. (2006) developed a portable device to present air samples to conditioned M. croceipes and to interpret the food-searching behavior of five conditioned wasps held in a cartridge. Video footage of the wasp behavior is recorded remotely in a laptop computer. An user-developed software program analyzes the video of wasps to determine when they have detected a chemical that they were conditioned to recognize (Utley et al., 2007). A result can be determined within 20-30 s after the wasps are exposed to the air sample. This device calls the Wasp Hound® (Rains et al., 2008).
40A different device has been developed to hold ten noctuid hawkmoths. With this device, the feeding response of each moth is measured by electromyography (King et al., 2004). A voltmeter is used to detect spikes in the signal from the feeding muscles of the moth. Five moths were conditioned to the target chemical, and five were used as an unconditioned control to determine if responses from the conditioned moths were false positives (King et al., 2004). However, this device is heavy (17 kg) and thus makes it less portable.
41These three portable structures make it ideal for testing in airports, subway stations, etc.
42It appears that free-moving insects are best suited to foraging in natural environment than restrained insects.
4. Conclusion
43Various biosensors are used in forensic sciences. However, mammalian biosensors have disadvantages unshared with those made with insects. Insects might be a preferred sensing method in scenarios that are deemed too dangerous to use mammalian. For example, canines that detect cadaver are often placed in dangerous conditions, either through exposure to toxic chemicals or unstable structures. Development of alternative sensing systems using insects would be advantageous because the loss of an insect sensor would be less expensive than losing a trained canine.
44Insects are not often used as biosensors for corpses detection. However, the necrophagous insects, i.e. those which colonize a corps, could be very effective as they are pledged with this particular ecosystem. They also show an array of instinctive and reflexive orienting behaviors in response to decaying flesh (King et al., 2004). The advantage of using instinctive behaviors of insects is that they are reflexive, occurring with a high degree of probability. Future detection systems might be developed around other insect species that have innate responses to chemical targets, or that have been selectively reared to have enhanced response to chemicals of interest. Parasitic wasps of fly pupae from carcasses seem to be highly successful to locate carcasses and hosts.
45Finally, further scientific research is needed to enhance the utility of insect-sniffer detection systems and, furthermore, to increase the level of acceptance from a larger community of research and commercial enterprises. Once this detection system has proven its ability to perform in a real-world commercial application, there should be a coincidental increase in interest to develop insect sensors for other attractive applications (Rains et al., 2008).
46Acknowledgements
47Christine Frederickx is financially supported by PhD grant from the Fonds pour la Formation à la Recherche dans l'Industrie et l'Agriculture (F.R.I.A.), Belgium.
Bibliographie
Amendt J. et al., 2000. Forensic entomology in Germany. Forensic Sci. Int., 113, 309-314.
Amendt J., Krettek R. & Zehner R., 2004. Forensic entomology. Naturwissenschaften, 91, 51-65.
Amine A., Mohammadi H., Bourais I. & Palleschi G., 2006. Enzyme inhibition-based biosensors for food safety and environmental monitoring. Biosens. Bioelectron., 21, 1405-1423.
APOPO, 2003. APOPO vapour detection technology, http://www.apopo.org, (03/13/10).
Badihi-Mossberg M., Buchner V. & Rishpon J., 2007. Electrochemical biosensors for pollutants in the environment. Electroanalysis, 19, 2015-2028.
Banaigs B., 2002. La communication chimique dans le monde vivant, http://perspectives.univ-perp.fr/spip.php?article14, (08/23/08).
Benecke M. & Lessig R., 2001. Child neglect and forensic entomology. Forensic Sci. Int., 120, 155-159.
Benecke M. & Wells D.J., 2001. DNA Techniques for forensic entomology. In: Castner J.H. & Byrd J.L., eds. Forensic entomology: the utility of arthropods in legal investigations. Boston, MA, USA: CRC Press, 341-352.
Bernstein D.A. & Nash P.W., 2008. Essentials of psychology. 4th ed. Boston, MA, USA: Wadsworth Publishing.
Bourel B. et al., 2001. Morphine extraction in necrophagous insects remains for determining ante-mortem opiate intoxication. Forensic Sci. Int., 120, 127-131.
Brossut R., 1996. Phéromones : la communication chimique chez les animaux. Paris : CNRS Éditions.
Carcaud J., Roussel E., Giurfa M. & Sandoz J.C., 2009. Odour aversion after olfactory conditioning of the sting extension reflex in honeybees. J. Exp. Biol., 212(5), 620-626.
Clark L.C. & Lyons C., 1962. Electrode systems for continuous monitoring in cardiovascular surgery. Ann. N.Y. Acad. Sci., 102, 29-45.
Cornaz J.N., 2008. La recherche de cadavres et de sang en milieu aquatique par des chiens. PolCant info, 72, 14-15.
Davis P.J., Wadhams L. & Baylis J.S., 2005. Detection of odors using insects. Patent UK US2005009444.
Gennard D.E., 2007. Forensic entomology: an introduction. Chichester, UK: John Wiley & Sons Ltd.
Given J.A., 2003. Controlled substance training aids for DoD. In: 3rd National Detector Dog Conference, May 19-23, 2003, North Miami Beach, Florida, USA.
Grassberger M. & Frank C., 2003. Temperature-related development of the parasitoid wasp Nasonia vitripennis as forensic indicator. Med. Vet. Entomol., 17, 257-262.
Gupta A. & Setia P., 2004. Forensic entomology: past, present and future. Aggrawal's Internet J. Forensic Med. Toxicol., 5(1), 50-53.
Hall R.D. & Huntington T.E., 2009. Introduction: perception and status of forensic entomology. In: Byrd J.H. & Castner J.L., eds. Forensic entomology: the utility of arthropods in legal investigations. Vol. 2. Boca Raton, FL, USA: CRC Press, 1-16.
Hammer M. & Menzel R., 1995. Learning and memory in the honeybee. J. Neurosci., 15(3), 1617-1630.
Harper R.J. & Furton K.G., 2007. Biological detection of explosives. In: Yinon J., ed. Counterterrorist detection techniques of explosives. Amsterdam, The Netherlands; Oxford, UK: Elsevier, 395-431.
Keil T.A., 1999. Morphology and development of the peripheral olfactory organs. In: Hansson B.S., ed. Insect olfaction. Berlin, Germany; New York, USA: Springer, 5-48.
King T.L., Horine F.M., Daly K.C. & Smith B.H., 2004. Explosives detection with hard-wired moths. IEEE Trans. Instrum. Meas., 53(4), 1113-1118.
Lanzoni M., Stagni C. & Riccò B., 2009. Smart sensors for fast biological analysis. Microelectron. J., 40, 1345-1349.
Lasseter A.E., Jacobi K.P., Farley R. & Hensel L., 2003. Cadaver dog and handler team capabilities in the recovery of buried human remains in the southeastern United States. J. Forensic Sci., 48(3), 617-621.
Leffingwell J.C., 2002. Olfaction-Update No 5, http://www.leffingwell.com/olfaction.htm, (03/12/10).
Meierhenrich U.J., Golebiowski J., Fernandez X. & Cabrol-Bass D., 2004. The molecular basis of olfactory chemoreception. Angew. Chem. Int. Ed., 43(47), 6410-6412.
Menzel R. & Muller U., 1996. Learning and memory in honeybees: from behavior to neural substrates. Annu. Rev. Neurosci., 19, 379-404.
Nolan R., Weinstein S. & Weinstein C., 1978. Electroencephalographic studies of specifically-conditioned explosives detecting rats. In: Proceedings of the new concepts symposium and workshop on detecting and identification of explosives, October 30-31 and November 1, Reston, Virginia, USA, 201-205.
Oesterhelweg L. et al., 2008. Cadaver dogs. A study on detection of contaminated carpet squares. Forensic Sci. Int., 174, 35-39.
Olson D.M. et al., 2003. Parasitic wasps learn and report diverse chemicals with unique conditionable behaviors. Chem. Senses, 28(6), 545-549.
Otto J., Brown M.F. & Long W., 2002. Training rats to search and alert on contraband odors. Appl. Anim. Behav. Sci., 77(3), 217-232.
Pavlov I. & Anrep G.V., 1960. Conditioned reflexes: an investigation of the physiological activity of the cerebral cortex. New York, USA: Dover Publications.
Rains G.C., Tomberlin J.K., D'Alessandro M. & Lewis W.J., 2004. Limits of volatile chemical detection of a parasitoid wasp, Microplitis croceipes, and an electronic nose: a comparative study. Trans. Asae, 47(6), 2145-2152.
Rains G.C., Utley S.L. & Lewis W.J., 2006. Behavioral monitoring of trained insects for chemical detection. Biotechnol. Progr., 22(1), 2-8.
Rains G.C., Tomberlin J.K. & Kulasiri D., 2008. Using insect sniffing devices for detection. Trends Biotechnol., 26(6), 288-294.
Rebmann A., David E. & Sorg M.H., 2000. Cadaver dog handbook: forensic training and tactics for the recovery of human remains. Boca Raton, FL, USA: CRC Press.
Ricciuti E., 2007. Science 101: forensics. New York, USA: HarperCollins Publishers.
Riley J.R. & Smith A.D., 2002. Design considerations for an harmonic radar to investigate the flight of insects at low altitude. Comput. Electron. Agric., 35(2-3), 151-169.
SARDUS, 1992. Search and rescue dogs of the United States, http://www.sardogsus.org/id13.html, (12/01/2011).
Scheiner R., Page R.E. & Erber J., 2001. The effects of genotype, foraging role, and sucrose responsiveness on the tactile learning performance of honey bees (Apis mellifera L.). Neurobiol. Learn. Memory, 76(2), 138-150.
Settles G.S. & Kester D.A., 2001. Aerodynamic sampling for landmine trace detection. In: Proceedings of SPIE—Detection and Remediation Technologies for Mines and Minelike Targets.
Seuter E.J., 2003. The dog days of detection, http://www.nobombs.net/dog_days_of_detections.html, (03/12/10).
Shaw J.A. et al., 2005. Polarization lidar measurements of honey bees in flight for locating land mines. Opt. Express, 13(15), 5853-5863.
Skipper B.F., 1938. Behavior of organisms: experimental analysis. New York, USA: D. Appleton-Century Company, Inc.
SPAWAR, 1992. U.S. Navy Marine Mammal Program, http://www.spawar.navy.mil/sandiego/technology/mammals/index.html, (03/10/10).
Stengl M., Ziegelberger G., Boekhoff I. & Krieger J., 1999. Perireceptor events and transduction mechanisms in insect olfaction. In: Hansson B.S., ed. Insect olfaction. Berlin, Germany; Heidelberg, Germany: Springer-Verlag, 49-66.
Stephens D.W., 1993. Learning and behavioural ecology: incomplete information and environmental predictability. In: Papaj D.R. & Lewis A.C., eds. Insect learning: ecological and evolutionary perspectives. New York, USA: Chapman and Hall, 195-218.
Takasu K., Rains G. & Lewis W., 2007. Comparison of detection ability of learned odors between males and females in the larval parasitoid Microplitis croceipes. Entomol. Exp. Appl., 122, 247-251.
Tertuliano M., Olson D., Rains G. & Lewis W., 2004. Influence of handling and conditioning protocol on learning and memory of Microplitis croceipes. Entomol. Exp. Appl., 110, 165-172.
Tokarskyy O. & Marshall D.L., 2008. Immunosensors for rapid detection of Escherichia coli O157:H7 - Perspectives for use in the meat processing industry. Food Microbiol., 25, 1-12.
Tomberlin J., Tertuliano M., Rains G. & Lewis W., 2005. Conditioned Microplitis croceipes Cresson (Hymenoptera: Braconidae) detect and respond to 2, 4-DNT: development of a biological sensor. J. Forensic Sci., 50(5), 1187-1190.
Tomberlin J., Rains G. & Sanford M., 2008. Development of Microplitis croceipes as a biological sensor. Entomol. Exp. Appl., 128, 249-257.
Turchetto M. & Vanin S., 2004. Forensic evaluations on a crime case with monospecific necrophagous fly population infected by two parasitoid species. Aggrawal's Int. J. Forensic Med. Toxicol., 5(1), 12-18.
Utley S.L., Rains G.C. & Lewis W., 2007. Behavioral monitoring of Microplitis croceipes, a parasitoid wasp, for detecting target odorants using a computer vision system. Trans. ASABE, 50(5), 1843-1849.
Voss S.C., Spafford H. & Dadour I.R., 2009. Hymenopteran parasitoids of forensic importance: host associations, seasonality, and prevalence of parasitoids of carrion flies in Western Australia. J. Med. Entomol., 46(5), 1210-1219.
Weetall H.H., 1996. Biosensor technology - What? Where? When? And Why? Biosens. Bioelectron., 11(1-2), R1-R4.
Weinstein S., Weinstein C. & Drozdenko R., 1992. The challenge of biodetection for screening persons carrying explosives. In: Proceedings of the 1st International Symposium on Explosive detection technology, November 13-15, 1991, Atlantic City International Airport, N.J., USA. Atlantic City International Airport, N.J., USA: FAA Technical Center.
Wyss C. & Cherix D., 2006. Traité d'entomologie forensique : les insectes sur la scène de crime. Lausanne, Suisse : Presses Polytechniques et Universitaires romandes.
Yinon J. & Zitrin S., 1996. Modern methods and applications in analysis of explosives. Chichester, UK; New York, USA: Wiley.
Pour citer cet article
A propos de : Christine Frederickx
Univ. Liege - Gembloux Agro-Bio Tech. Department of Functional and Evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium). E-mail: cfrederickx@student.ulg.ac.be
A propos de : François J. Verheggen
Univ. Liege - Gembloux Agro-Bio Tech. Department of Functional and Evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
A propos de : Éric Haubruge
Univ. Liege - Gembloux Agro-Bio Tech. Department of Functional and Evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).