


- Accueil
- Volume 14 (2010)
- numéro 4
- Relation entre la composition floristique et des indicateurs de la fragmentation du paysage dans une région de transition forêt-savane ivoirienne


Visualisation(s): 3238 (47 ULiège)
Téléchargement(s): 302 (8 ULiège)

Relation entre la composition floristique et des indicateurs de la fragmentation du paysage dans une région de transition forêt-savane ivoirienne

Notes de la rédaction
Reçu le 1 juillet 2009, accepté le 2 mars 2010
Résumé
La fragmentation du paysage forestier est connue comme étant un facteur important de la dégradation des forêts. Dans cet article, nous analysons le cas de la zone de contact forêt-savane à l'Est de la Côte d'Ivoire, où les savanes progressent rapidement au détriment des forêts. À l'aide d'inventaires forestiers et des propriétés spectrales et spatiales d'une image Landsat ETM+, nous avons établi l'existence de deux gradients floristiques indépendants au sein des forêts, pouvant être mis en relation avec différents aspects de la fragmentation. Le gradient principal est corrélé au degré d'ouverture des peuplements et à l'abondance des espèces héliophiles. Sur le plan spectral, il se traduit par une variation nette dans les bandes du rouge et du proche infrarouge. En termes de configuration paysagère, ce gradient floristique est significativement corrélé à la taille et à l'isolement des ilots forestiers. Un second gradient floristique est corrélé au périmètre des fragments forestiers et pourrait donc refléter l'effet des lisières. Ces résultats permettent d'envisager une approche opérationnelle à partir de données multispectrales de résolution moyenne et d'indices de configuration spatiale pour l'évaluation et le suivi opérationnel des dégradations forestières à l'échelle régionale.
Abstract
Relation between floristic composition and landscape fragmentation indicators in a transition forest-savanna area of Côte d'Ivoire. Forest landscape fragmentation is known to be a significant factor of forest degradation. In this article, we analyze the case of forest-savanna contact of East Côte d'Ivoire, where savannas progress quickly to the detriment of forests. Using forest inventories and the spectral and spatial properties of a Landsat ETM+ image, we established the existence of two independent floristic gradients within forests, which were correlated to various aspects of forest fragmentation. The main floristic gradient was correlated with the degree of opening of forest stands and with the abundance of heliophilics. Spectrally, this gradient clearly correlates with a variation in the red and near infra-red bands. In terms of landscape structure, it is significantly related on the size and the isolation of small forest patches. A second floristic gradient is correlated to the perimeter of forest fragments, and may thus reflect an edge effect. These results make it possible to consider an operational approach based on multispectral data of average resolution and pattern metrics for operational assessment and monitoring of forest degradations on regional scales.
Table des matières
1. Introduction
1L'occupation du sol dans les pays tropicaux change rapidement (Veldkamp et al., 2001 ; Hansen et al., 2008) et la fragmentation des habitats naturels liée aux activités humaines est devenue l'un des processus dominants de la dynamique paysagère. Par la réduction de la taille des fragments et l'augmentation de leur isolement (Fahrig, 2003 ; Bogaert et al., 2004), la fragmentation modifie la dynamique des forêts tropicales et altère les cycles de reproduction des espèces (Aguilar et al., 2006 ; Lindenmayer et al., 2006). La fragmentation entraine également une augmentation de la surface des lisières (Laurance et al., 1997 ; Hennenberg et al., 2008) et des trouées (Asner et al., 2004). Elle modifie ainsi le microclimat au sein des fragments forestiers par une baisse de l'humidité et une augmentation de l'éclairement (Heithecker et al., 2007 ; Laurance et al., 2008), ce qui stimule la prolifération des héliophytes invasives comme Chromolaena odorata (L.) R.M.King & H.Rob. Ce microclimat augmente la sensibilité des fragments forestiers au feu (Cochrane, 2003).
2En somme, la fragmentation des habitats naturels peut donc induire des modifications de la configuration forestière (réduction de taille, isolement des fragments, etc.) et de la structure forestière (densité en tiges, surface terrière) des fragments (Freitas et al., 2005 ; Collins et al., 2009) ainsi que de leur composition floristique (Cabacinha et al., 2009 ; Munro et al., 2009).
3Pour mettre en place un suivi opérationnel de la fragmentation et de la déforestation en milieu tropical, il importe d'utiliser des approches peu complexes mais robustes, facilement reproductibles, pouvant couvrir de grandes superficies et permettant d'étudier des dynamiques temporelles avec suffisamment de recul et de répétitivité (DeFries et al., 2007). L'utilisation de données de télédétection optique multispectrale de résolution moyenne (exemple Landsat TM, Landsat ETM à 30 m de résolution ou Aster à 15 m de résolution) permet de relever ce défi (Schowengerdt, 2007 ; Lillesand et al., 2008). En effet, à partir de l'information spectrale, ces données permettent de différencier des types forestiers relativement à leur état de dégradation (Asner et al., 2005 ; DeFries et al., 2007), même si l'on note une saturation du signal spectral pour des valeurs élevées de biomasse (supérieures à 250 t.ha-1) (Foody et al., 2003). À partir de l'information spatiale issue de ces mêmes données de télédétection, notamment via des indices de configuration, il est également possible de quantifier l'organisation dans l'espace des fragments forestiers (McGarigal et al., 2002 ; Barima et al., 2009).
4Dans cet article, nous cherchons à mettre en évidence une relation entre ces deux types d'information (spectrale et spatiale) et la structure et la composition forestière des fragments sur un front de déforestation dans l'Est de la Côte d'Ivoire. Par structure, nous entendons la densité des tiges ou leur surface terrière ; la composition forestière correspond aux différentes informations tirées de la liste floristique (gradient floristique, diversité et comportement des espèces vis-à-vis de la lumière). Nous tentons de répondre aux questions suivantes :
5– peut-on établir un lien entre la fragmentation et la structure forestière estimée, soit à partir de la radiométrie, soit par la mesure sur le terrain de la densité des arbres ?
6– les différents aspects de la fragmentation (isolement, taille, lisière) induisent-ils des effets contrastés sur la composition forestière des fragments ?
2. Méthodes
2.1. Zone d'étude
7Les travaux ont été effectués dans la zone de transition forêt-savane ivoirienne, dans une région située entre 03°00' - 03°24' de longitude Ouest et 07°41' - 08°04' de latitude Nord. Elle est localisée au sein du département de Tanda à l'Est de la Côte d'Ivoire (Figure 1). Le climat de la région est du type tropical à tendance sub-humide (Guillaumet et al., 1971). Il est caractérisé par une forte irrégularité de la pluviométrie (10 mm - 160 mm par mois), un déficit hydrique annuel compris entre 400 et 450 mm et une saison sèche qui couvre la période allant de novembre à février. La température moyenne annuelle est de 26 °C. Le relief présente un gradient altitudinal qui passe progressivement de 200 m dans le Sud-Ouest à 700 m dans le Nord-Est de la zone d'étude (Avenard, 1971). Les sols présentent une alternance de sols ferrallitiques moyennement et fortement désaturés dans l'horizon B (Perreaud, 1971). La région se trouve dans le secteur mésophile du domaine guinéen avec pour climax dominant la forêt dense humide semi-décidue. Guillaumet et al. (1971) assimilent le groupement végétal de la région d'étude au type à Aubrevillea kerstingii (Harms) Pellegr. et Khaya grandifolia C.DC. Dans le détail, la végétation est répartie en trois types de formations :
8– les forêts, dont la forêt secondaire à Musanga, sont disséminées dans toute la région, hormis dans la partie Sud-Ouest ; la forêt à Musanga présente souvent l'aspect d'une forêt bien conservée remaniée à Ceiba pentandra (L.) Gaertn. et Khaya grandifolia,
9– la savane arborée à rôniers (Borassus aethiopum Mart.), présentant une strate arbustive, est localisée dans l'Est et à l'Ouest de la région ; cette savane est entourée par endroits de forêts denses donnant l'aspect de savane incluse,
10- la savane arbustive, dominée par les Mimosaceae à strate herbeuse riche en Imperata cylindrica (L.) Raeusch, est répartie dans toutes les parties de la région.
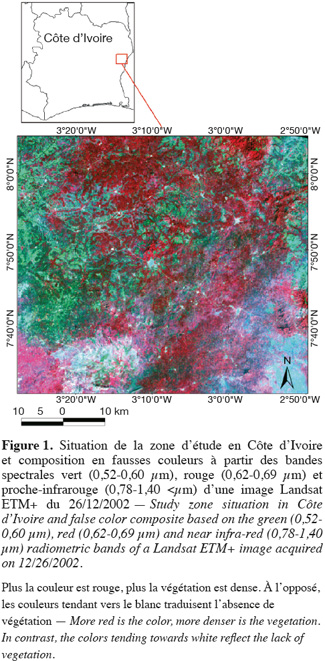
11Dans cette région, le taux de déforestation a atteint en 2000, 4 % par an (Barima et al., 2009), taux supérieur au taux national qui était de 2,9 % par an selon Achard et al. (2002) et 3,1 % par an selon la FAO (FAO, 2001). Il en résulte des surfaces de lisières plus importantes et une hétérogénéité accrue du milieu (Rudel et al., 2005 ; Hennenberg et al., 2008) dans cette région de contact forêt-savane, en comparaison aux autres régions du pays.
2.2. Classification radiométrique semi-supervisée
12Afin de cartographier les zones forestières de la région d'étude, une classification semi-supervisée a été réalisée sur une image Landsat ETM+ (Enhanced Thematic Mapper plus) datant du 26/12/2002 (Figure 1). Cette image, de 30 m de résolution, a d'abord été orthorectifiée sur base des coordonnées géographiques de points d'amer localisés sur le terrain au GPS (croisement des grands axes routiers, grands bâtiments, affleurements rocheux). La précision géométrique du calage entre ces points et l'image Landsat ETM+ était inférieure à un pixel (30 m). La classification s'est faite en trois étapes.
13D'abord, une classification non supervisée sur base des bandes vert (0,52-0,60 µm), rouge (R : 0,62-0,69 μm) et proche-infrarouge (PIR : 0,78-1,40 μm) a été effectuée par maximum de vraisemblance (Rogan et al., 2004 ; Lillesand et al., 2008) et a permis d'obtenir 12 classes d'occupation du sol. Cet algorithme se base sur une analyse statistique de la distribution des vecteurs spectraux de l'échantillon pour définir des zones de probabilité équivalente autour des centres. La probabilité d'appartenance à chacune des classes est calculée pour chaque vecteur spectral et le vecteur est affecté à la classe pour laquelle la probabilité est la plus élevée. Un avantage considérable de cette méthode est qu'elle fournit pour chaque pixel, en plus de la classe à laquelle il a été affecté, un indice de certitude lié à ce choix (Schowengerdt, 2007 ; Lillesand et al., 2008). Ensuite, dans une deuxième étape, un regroupement thématique des 12 classes obtenues en fonction du type d'occupation du sol (forêts, savanes, champs, jachères, sol nu) a été réalisé de manière visuelle. Cette étape a permis de fusionner des classes radiométriques et thématiques similaires. En effet, les 12 classes obtenues lors de la première étape présentaient une forte hétérogénéité et offraient trop de détails au sein d'une même classe thématique. Huit classes d'occupation du sol, dont trois classes de forêt, quatre classes de formations agricoles et une classe de sols nus et d'habitations ont ainsi été obtenues. Enfin, dans une dernière étape, les statistiques radiométriques des 8 classes ont été extraites des bandes spectrales du R et du PIR et projetées sur des axes orthonormés afin de regrouper les classes radiométriquement proches. Ces canaux sont les plus employés pour les études de végétation car ils permettent de mieux distinguer les différents types de végétation de strates et de densités variables (Lillesand et al., 2008 ; Barima et al., 2009). Les étapes de traitement et de classification d'image ont été réalisées dans le logiciel ENVI 4.2.
14La classification effectuée a été validée après vérification par reconnaissance qualitative des classes d'occupation du sol sur le terrain, à partir de 200 sites de référence se trouvant dans des zones homogènes de taille supérieure ou égale à 20 pixels. Un coefficient de kappa = 80,50 % a été obtenu ; selon Pontius (2000) et Oloukoi et al. (2006), une étude de l'occupation du sol peut être validée si le coefficient kappa est supérieur à 50 ou 75 %, selon les cas.
2.3. Inventaires forestiers
15Quarante six sites d'inventaires forestiers ont été définis dans les classes forestières à partir d'un échantillonnage aléatoire, stratifié (forêt, non-forêt) sur la base de la classification de l'image Landsat ETM+ 2002. Ces inventaires forestiers ont été effectués suivant la méthode préconisée par Senterre (2005) : les cent premiers individus de diamètre à hauteur de poitrine ≥ 10 cm (Condit et al., 1996 ; Senterre, 2005) rencontrés à partir du centre du site ont été recensés (espèce) et mesurés. La surface dans laquelle ces cents individus ont été inventoriés a été déterminée avec une boussole et un télémètre laser, ce qui a permis de calculer la densité (d) du peuplement pour chaque site d'inventaire. La nomenclature des spécimens botaniques a suivi celle de Lebrun et al. (1991-1997). Les espèces ont ensuite été regroupées suivant leurs stratégies écologiques par rapport à la lumière en héliophytes et en non-héliophytes (Molino et al., 2001 ; Arim et al., 2006) sur base des travaux de Hawthorne (1996) et de Bakayoko (2005). Le pourcentage d'espèces héliophiles (h) a ainsi été déterminé dans chaque site d'inventaire forestier.
2.4 Détermination de la configuration du paysage
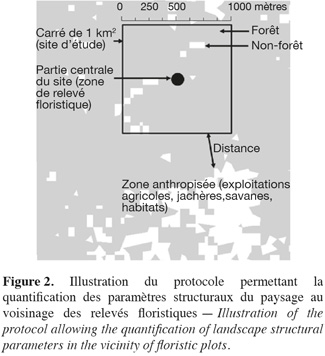
16Un carré de 1 km2 (1 111 pixels environ) a été délimité sur l'image Landsat autour du centre de chacun des sites d'inventaire forestier (Figure 2). Le nombre de taches (N), le périmètre (P) et l'aire des forêts (A) ont été déterminés dans chaque carré à l'aide des logiciels Fragstats 3.3 et ArcGis 9.2. Dans l'optique d'identifier l'influence anthropique sur la composition floristique des relevés et la configuration des forêts, la distance (D) entre un carré et la classe anthropisée la plus proche a été déterminée. Pour cela, nous avons considéré comme espaces anthropisés les classes non forestières (exploitations agricoles, savanes, jachères et habitats anthropogènes) dont les surfaces étaient supérieures à 1 pixel (900 m2) afin d'éviter des confusions avec des phénomènes naturels tels que les chablis. Nous supposons en effet que les chablis ont des tailles de 900 m2 au maximum (Webster et al., 2005) et qu'ils causent des perturbations d'intensité minime.
2.5. Analyse des données
17Une ordination des sites d'inventaire a été faite sur base de leur composition floristique. L'analyse des correspondances redressée (ACR ou Detrended Correspondence Analysis) a été préférée à l'analyse canonique afin de supprimer l'effet d'arche (Legendre et al., 1998) que nous avons observé en effectuant l'analyse canonique. L'ACR reste à ce jour la principale méthode d'analyse indirecte de gradient, c'est-à-dire d'analyse de la variance d'un lot de données floristiques correspondant à des relevés placés le long d'un gradient environnemental inconnu a priori (Legendre et al., 1998). Un diagramme de double projection (Jongman et al., 1995) a été construit afin de tester la relation linéaire entre les coordonnées des sites d'inventaires sur les axes de l'ACR et les valeurs de densité (d), les stratégies écologiques (pourcentage d'espèces héliophiles par relevé : h) et les indices de configuration paysagère (P, A, N et D). Ces analyses ont été effectuées à partir du logiciel CANOCO 4.5. Des régressions linéaires simples (c'est-à-dire à un facteur) et multiples ont également été effectuées suivant la stratégie préconisée par Grafen et al. (2008). La significativité des variables explicatives dans les modèles de régression multiple a été testée sur base des sommes des carrés ajustées (SSadj). Cette approche rend les tests indépendants de l'ordre dans lequel les variables sont introduites dans le modèle, par opposition à la somme des carrés séquentielle (Grafen et al., 2008). En effet, SSadj indique pour chaque variable explicative le surcroit de variance expliquée une fois que les autres variables ont été prises en compte. Cette approche permet d'identifier la proportion de variance expliquée indépendamment par chaque variable, par rapport à celle partagée avec d'autres variables.
18Les espèces indicatrices ont été identifiées à partir d'une classification des inventaires suivant la méthode IndVal (Dufrêne et al., 1997) avec un seuil de significativité de < 1 %. Cette méthode consiste à identifier les espèces indicatrices à l'aide d'un test de permutation qui combine la fidélité des espèces (proportion de sites appartenant à un groupe où l'espèce est présente) et leur spécificité (à quel point une espèce ne se trouve que dans le groupe considéré) (Dufrêne et al., 1997).
3. Résultats
3.1. Classification radiométrique
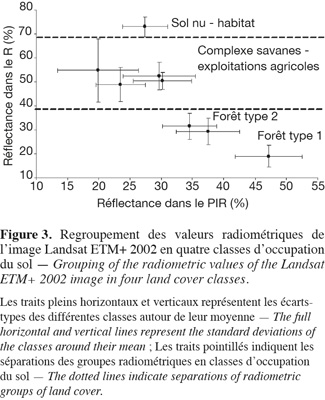
19L'extraction des statistiques radiométriques des bandes spectrales R et PIR des classes créées suite à une classification non supervisée de l'image Landsat ETM+ a permis un regroupement en quatre classes distinctes (Figure 3). Il s'agit d'une classe de sol nu-habitat (anthropogénique), un complexe « savanes-exploitations agricoles » (composés de savanes herbeuses, de savanes boisées, d'exploitations agricoles et de jachères) et de deux types de forêts. Dans l'ensemble, les classes de sol nu-habitat et les forêts se distinguent clairement du nuage formé par la classe hétérogène (complexe savanes-exploitations agricoles). Par contre, les savanes boisées, les savanes herbeuses, les exploitations agricoles et les jachères sont difficilement séparables du fait de leur ressemblance spectrale. Pour ce qui est de la classe forestière, on peut distinguer un gradient allant des taux de réflectance les plus faibles dans le PIR (34,53 %) aux plus élevées (47,19 %) (Figure 3). Deux types de forêts (type 1 et type 2), séparables sur base radiométrique, semblent exister dans la zone d'étude.
3.2. Composition floristique
20Les inventaires ont permis de comptabiliser au total 4 600 arbres répartis en 186 espèces, 124 genres et 37 familles. Les individus les plus abondants appartiennent à la famille des Meliaceae (22,08 % des individus) et des Moraceae (17,13 % des individus) respectivement dans les forêts de type 1 et de type 2. Les principales espèces indicatrices (avec un seuil de significativité < 0,01) de la forêt de type 1 sont : Cola millenii K.Schum., Trichilia prieuriana A.Juss., Celtis wightii Planch., Chrysophyllum giganteum A.Chev. et Napoleonaea vogelii Hook. & Planch. Celles de la forêt de type 2 sont : Anthocleista djalonensis A.Chev., Holarrhena floribunda (G.Don) T.Durand & Schinz, Ficus exasperata Vahl et Bombax buonopozense P.Beauv.
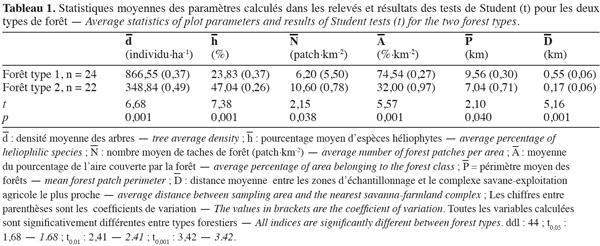
21Les résultats des variables floristiques étudiées (d, h) pour chaque type de forêt ainsi que les résultats des tests de comparaison de moyennes (test de Student) (Tableau 1) montrent que les relevés effectués dans les forêts de type 1 sont caractérisés par une densité moyenne relativement élevée en arbres (866,55 individus.ha-1) par rapport aux relevés effectués dans le deuxième type de forêt (348,84 individus.ha-1). Ces derniers possèdent par contre des taux importants d'héliophytes (47,04 %), quand seules 23,83 % des espèces recensées dans les forêts de type 1 appartiennent à ce groupe.
22Les gradients floristiques présents dans le jeu d'inventaires forestiers ont été caractérisés par une ACR. Les deux premiers axes canoniques qui représentent 60,15 % de la variabilité totale (axe 1 = 41,37 % ; axe 2 = 18,78 %) expliquent la majeure partie de la variabilité du jeu de données. La projection des scores des relevés sur ces axes permet d'obtenir une visualisation des résultats de l'ACR en deux dimensions (Figure 4). Dans le diagramme d'ordination ainsi produit, les deux principaux types radiométriques de forêts se séparent assez bien, les relevés des forêts de type 2 possédant des valeurs plus élevées sur le premier axe. Le test de Student effectué avec les scores des relevés sur l'axe principal de l'ACR montre qu'il existe une différence significative entre les moyennes de chaque type de forêt (p < 0,001).
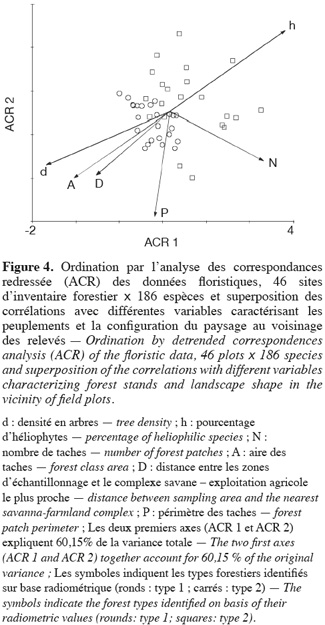
3.3. Configuration du paysage
23Au niveau de la configuration du paysage, la forêt de type 2 se caractérise en moyenne par un nombre de taches plus important, soit 10,60 taches par km2 contre 6,20 taches par km2 pour les forêts de type 1 (p < 0,05) (Tableau 1). Le fait que la proportion de la classe forêt soit moins importante pour le type 2 que pour le type 1 nous permet de dire que les forêts de type 2 sont plus fragmentées (taches nombreuses et petites) que les forêts de type 1. De plus, leur proximité aux zones anthropisées (171,70 m contre 552,54 m pour la forêt de type 1, p < 0,001) indique très probablement que les forêts de type 2 sont fortement influencées par les activités humaines.
3.4. Déterminisme des gradients floristiques
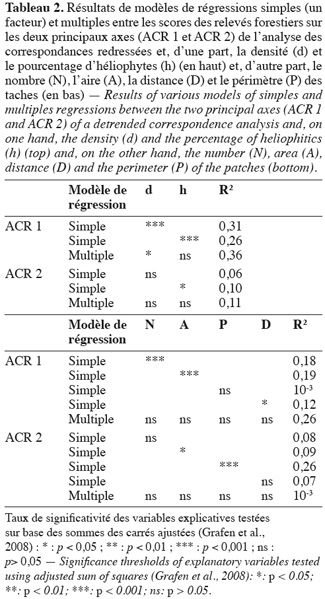
24Pour pouvoir interpréter les gradients floristiques mis en évidence, un diagramme de double projection est présenté montrant la relation linéaire entre les deux axes principaux de l'ACR et les valeurs de densité et de stratégie écologique d'une part, et d'autre part les données relatives à la configuration paysagère (P, A, N et D) (Figure 4). Le score de ces variables sur les deux axes correspond à la pente de la régression et donc la direction des flèches peut être traduite comme la direction de variation maximale de chaque variable. Ce résultat peut être approfondi au travers de régressions linéaires simples ou multiples (Tableau 2). Il apparait que l'axe 1 est significativement corrélé à la densité en tiges (R2 = 0,31 ; p < 0,001) et au taux d'héliophytes (R2 = 0,26 ; p < 0,001). Un modèle plus complet de régression multiple impliquant ces deux variables ensemble ne produit pas un gain sensible de variance expliquée (R2 = 0,36) et les variables ne sont plus que faiblement ou non significatives selon le test basé sur le SSadj. On peut donc en déduire que ces deux variables sont redondantes. Seul le taux d'héliophytes est corrélé (faiblement) (R2 = 0,10 ; p < 0,05) avec l'axe 2 de l'ACR. Pour ce qui est des paramètres structuraux, l'axe 1 est très significativement corrélé à N (R2 = 0,18 ; p < 0,01) et A (R2 = 0,19 ; p < 0,01) et significativement corrélé à D (R2 = 0,12 ; p < 0,05), mais pas à P (R2 = 10-3 ; p > 0,05). Notons qu'ici encore, la régression multiple permet de détecter une certaine redondance entre les variables, puisqu'elles perdent leur significativité une fois prises ensemble. L'axe 2 de l'ACR est quant à lui corrélé très significativement avec P (R2 = 0,26 ; p < 0,001) et faiblement avec D (R2 = 0,12 ; p < 0,05). En somme, l'axe 1 est un gradient floristique simultanément corrélé au pourcentage d'héliophytes, à la densité en tiges et à la taille des fragments forestiers. L'axe 2 est un axe floristiquement indépendant du premier et est corrélé au périmètre. Ce second axe reflète donc vraisemblablement des effets induits par les lisières. La distance aux zones anthropisées ne semble pas être une variable explicative de la composition floristique des forêts de types 1 et 2 pris ensemble. Cependant, sa faible corrélation avec l'axe 1 de l'ACR (R2 = 0,12 ; p < 0,05) pourrait suggérer qu'une forêt proche d'une zone anthropisée a un risque élevé d'ouverture et de perturbation.
4. Discussion
25Cette étude confirme que l'utilisation de données multispectrales de résolution moyenne (par exemple Landsat) permet de quantifier les dégradations intervenues dans le paysage forestier (Asner et al., 2005 ; DeFries et al., 2007). Une approche reproductible, basée sur deux canaux spectraux, le R et le PIR, permet en effet la distinction de sous-types forestiers pertinents en termes de densité, de composition, de stratégie dominante et de niveau de dégradation. De plus, l'analyse de l'occupation du sol de la zone d'étude du début des années 1980 jusqu'aux années 2000 (Barima et al., 2009) a montré que la plupart des surfaces occupées par les forêts dégradées actuellement étaient initialement des forêts de type 1, c'est-à-dire une forêt de réflectance maximale dans le PIR, de densité en arbres et de surface plus élevées que les forêts de type 2. Les forêts de la zone d'étude ont donc subi des dégradations d'intensités diverses. Ce fait est confirmé par les corrélations significatives mises en évidence entre le gradient floristique et des indicateurs d'ouverture du couvert (héliophytes, densité).
26Nous avons également mis en évidence une corrélation nette entre la dégradation des forêts (changement de couverture de la canopée observée via la télédétection) et la fragmentation du milieu forestier, aisément quantifiable à l'aide d'indices spatiaux basés sur le nombre de taches et la proportion de forêts dans le paysage. Cette fragmentation, due à l'action de l'homme, semble produire deux effets indépendants sur la composition des forêts, qui se traduisent par les deux gradients floristiques orthogonaux que nous avons observés. Ainsi on observe d'une part, un effet de la taille et de l'isolement des fragments (mesurés par N et A). On en conclut que des fragments plus petits et isolés ont tendance à être moins denses en arbres et plus riches en espèces héliophiles conformément aux observations de Bakayoko (2005) en Côte d'Ivoire et de Tabarelli et al. (2008) au Brésil. Cet effet pourrait s'expliquer par le fait que les forêts les plus fragmentées sont nécessairement les plus perturbées par les activités humaines qui ont conduit à la déforestation aux alentours (coupes, débardages, collecte, activités agricoles, etc.) (Lindenmayer et al., 2006).
27À ce premier gradient floristique se superpose un second, qui est lui vraisemblablement lié à l'effet des lisières forestières dont les répercussions sur les fragments de forêts sont nombreux et complexes (Murcia, 1995 ; Gascon et al., 2000). La lisière permet en effet à la lumière et au vent de pénétrer latéralement dans le fragment de forêt, ce qui entraine un changement de microclimat le long d'une large bande de la forêt adjacente (Laurance et al., 2008). Ces changements affectent la configuration des forêts, la distribution des espèces animales et végétales et augmente considérablement la mortalité des arbres près des lisières (Laurance, 2006 ; Tabarelli et al., 2008). Les effets des lisières peuvent se faire sentir à des distances importantes à l'intérieur du fragment forestier allant de 100-300 m jusqu'à 1 000 m (Murcia, 1995 ; Laurance et al., 1997). Cela signifie que des fragments de 1 000 ha pourraient entièrement être des habitats de lisière. Le microclimat des lisières des fragments forestiers est également propice à la pénétration du feu, qui est une des causes notables de la dégradation des forêts tropicales (Cochrane, 2003). Le feu entrave la régénération le long des lisières en détruisant les réserves de semences enfouies dans le sol et compromet donc le processus de succession de la végétation (Vilà-Cabrera et al., 2008). Il induit une forte mortalité des essences forestières et favorise l'installation d'espèces pionnières ou même d'espèces exotiques ou rudérales (Tabarelli et al., 2008).
28Nos résultats, basés sur une méthodologie facilement reproductible et robuste, permettent d'envisager à large échelle la quantification des dégradations forestières, autant en termes de structure forestière que de composition. La faible résolution spatiale de capteur Landsat (30 m) relativement à la taille des arbres est compensée par l'information spectrale, suffisante pour distinguer des types forestiers relativement dégradés. Notons cependant qu'une saturation de l'information spectrale est signalée dans le cas des forêts multistrates les plus denses (biomasse > 250 t.ha-1) (Foody et al., 2003). Une information complémentaire, bien qu'indirecte, est cependant accessible au travers de la configuration spatiale du paysage. Le périmètre des taches, en particulier, permet de prédire des gradients floristiques différents de ceux accessibles par la simple radiométrie. De plus, dans une région où aucune réserve forestière n'existe et où le front forestier régresse ou se dégrade rapidement, notre approche permet de poser les bases d'une politique rationnelle de protection des forêts, en établissant des seuils minimaux de superficie des fragments, nécessaires à la préservation du biotope originel.
29Remerciements
30Cette recherche a fait l’objet d’un soutien de la Fondation Internationale pour la Science (Suède) par l’intermédiaire d’une bourse de recherche accordée à M. Y.S.S. Barima. Nous remercions les autorités du Conseil Général de Tanda (Côte d’Ivoire) pour les facilités qu’elles nous ont accordées pour accéder aux différents sites d’études.
Bibliographie
Achard F. et al., 2002. Determination of deforestation rates of the world's humid tropical forests. Science, 297(5583), 999-1002.
Aguilar R., Ashworth L., Galetto L. & Aizen M.A., 2006. Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecol. Lett., 9(8), 968-980.
Arim M. et al., 2006. Spread dynamics of invasive species. Proc. Natl Acad. Sci. USA, 103(2), 374-378.
Asner G.P. et al., 2004. Canopy damage and recovery after selective logging in Amazonia: field and satellite studies. Ecol. Appl., 14(4), S280-S298.
Asner G.P. et al., 2005. Selective logging in the Brazilian Amazon. Science, 310(5747), 480-482.
Avenard J.-M., 1971. Aspects de la géomorphologie. In: Avenard J.-M. et al., eds. Le milieu naturel de la Côte d'Ivoire. Paris : Orstom, 7-71.
Bakayoko A., 2005. Influence de la fragmentation forestière sur la composition floristique et la structure végétale dans le Sud-Ouest de la Côte d'Ivoire. Thèse de doctorat : Université de Cocody-Abidjan (Côte d'Ivoire).
Barima Y.S.S. et al., 2009. Dynamique paysagère en milieu de transition forêt-savane ivoirienne. Bois Forêts Trop., 299(1), 15-25.
Bogaert J., Ceulemans R. & Salvador-Van Eysenrode D., 2004. Decision tree algorithm for detection of spatial processes in landscape transformation. Environ. Manage., 33(1), 62-73.
Cabacinha C.D. & de Castro S.S., 2009. Relationships between floristic diversity and vegetation indices, forest structure and landscape metrics of fragments in Brazilian Cerrado. For. Ecol. Manage., 257(10), 2157-2165.
Cochrane M.A., 2003. Fire science for rainforests. Nature, 421(6926), 913-919.
Collins C.D., Holt R.D. & Foster B.L., 2009. Patch size effects on plant species decline in an experimentally fragmented landscape. Ecology, 90(9), 2577-2588.
Condit R. et al., 1996. Species-area and species-individual relationships for tropical trees: a comparison of three 50-ha plots. J. Ecol., 84(4), 549-562.
DeFries R. et al., 2007. Earth observations for estimating greenhouse gas emissions from deforestation in developing countries. Environ. Sci. Policy, 10(4), 385-394.
Dufrêne M. & Legendre P., 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr., 67(3), 345-366.
Fahrig L., 2003. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst., 34, 487-515.
FAO, 2001. Évaluation des ressources forestières mondiales en 2000. Rome : FAO.
Foody G.M., Boyd D.S. & Cutler M.E.J., 2003. Predictive relations of tropical forest biomass from Landsat TM data and their transferability between regions. Remote Sens. Environ., 85(4), 463-474.
Freitas S.R., Mello M.C.S. & Cruz C.B.M., 2005. Relationships between forest structure and vegetation indices in Atlantic rainforest. For. Ecol. Manage., 218(1-3), 353-362.
Gascon C., Williamson G.B. & da Fonseca G.A.B., 2000. Ecology - receding forest edges and vanishing reserves. Science, 288(5470), 1356-1358.
Grafen A. & Hails R., 2008. Modern statistics for the life sciences. Oxford, UK: Oxford University Press.
Guillaumet J.-L. & Adjanohoun E., 1971. La végétation de la Côte d'Ivoire. In: Avenard J.-M. et al., eds. Le milieu naturel de la Côte d'lvoire. Paris : Orstom, 157-263.
Hansen M.C. et al., 2008. Humid tropical forest clearing from 2000 to 2005 quantified by using multitemporal and multiresolution remotely sensed data. Proc. Natl Acad. Sci. USA, 105(27), 9439-9444.
Hawthorne W.D., 1996. Holes and the sums of parts in Ghanaian forest regeneration, scale and sustainable use. Edinburg, Scotland: Royal Society of Edinburg, 75-176.
Heithecker T.D. & Halpern C.B., 2007. Edge-related gradients in microclimate in forest aggregates following structural retention harvests in western Washington. For. Ecol. Manage., 248(3), 163-173.
Hennenberg K.J. et al., 2008. Detection of seasonal variability in microclimatic borders and ecotones between forest and savanna. Basic Appl. Ecol., 9(3), 275-285.
Jongman R.H.G., Ter Braak C.J.F. & van Tongeren O.F.R., 1995. Data analysis in community and landscape ecology. Cambridge, UK: Cambridge University Press.
Laurance W.F., 2006. Diversity in tropical forests. Science, 311(5766), 1376.
Laurance W.F. et al., 1997. Biomass collapse in Amazonian forest fragments. Science, 278(5340), 1117-1118.
Laurance W.F. & Curran T.J., 2008. Impacts of wind disturbance on fragmented tropical forests: a review and synthesis. Austral Ecol., 33(4), 399-408.
Lebrun J.P. & Stork A.L., 1991-1997. Énumération des plantes à fleurs d'Afrique tropicale. Genève, Suisse : Conservatoire et Jardin Botanique de la ville de Genève.
Legendre P. & Legendre L., 1998. Numerical ecology. Amsterdam; Oxford, UK: Elsevier.
Lillesand T.M., Kiefer R.W. & Chipman J.W., 2008. Remote sensing and image interpretation. Hoboken, NJ, USA: Wiley.
Lindenmayer D.B. & Fischer J., 2006. Habitat fragmentation and landscape change. An ecological and conservation synthesis. Washington, DC, USA: Island Press.
McGarigal K. & Cushman S.A., 2002. Comparative evaluation of experimental approaches to the study of habitat fragmentation effects. Ecol. Appl., 12(2), 335-345.
Molino J.F. & Sabatier D., 2001. Tree diversity in tropical rain forests: a validation of the intermediate disturbance hypothesis. Science, 294(5547), 1702-1704.
Munro N.T., Fischer J., Wood J. & Lindenmayer D.B., 2009. Revegetation in agricultural areas: the development of structural complexity and floristic diversity. Ecol. Appl., 19(5), 1197-1210.
Murcia C., 1995. Edge effects in fragmented forests. Implications for conservation. Trends Ecol. Evol., 10(2), 58-62.
Oloukoi J., Mama V.J. & Agbo F.B., 2006. Modélisation de la dynamique de l'occupation des terres dans le département des collines au Bénin. Télédétection, 6(4), 305-323.
Perreaud A., 1971. Le sol. In: Avenard J.M. et al., eds. Le milieu naturel de la Côte d'Ivoire. Paris : Orstom, 320-391.
Pontius R.G., 2000. Quantification error versus location error in comparison of categorical maps. Photogramm. Eng. Remote Sens., 66(8), 1011-1016.
Rogan J. & Chen D.M., 2004. Remote sensing technology for mapping and monitoring land-cover and land-use change. Prog. Plann., 61, 301-325.
Rudel T.K. et al., 2005. Forest transitions: towards a global understanding of land use change. Global Environ. Change, 15(1), 23-31.
Schowengerdt R.A., 2007. Remote sensing: models and methods for image processing. Oxford, UK: Elsevier/Academic Press.
Senterre B., 2005. Recherches méthodologiques pour la typologie de la végétation et la phytogéographie des forêts denses d'Afrique tropicale. Thèse de doctorat : Université Libre de Bruxelles (Belgique).
Tabarelli M., Lopes A.V. & Peres C.A., 2008. Edge-effects drive tropical forest fragments towards an early-successional system. Biotropica, 40(6), 657-661.
Veldkamp A. & Lambin E.F., 2001. Predicting land-use change. Agric. Ecosyst. Environ., 85(1-3), 1-6.
Vilà-Cabrera A., Saura-Mas S. & Lloret F., 2008. Effects of fire frequency on species composition in a Mediterranean shrubland. Ecoscience, 15(4), 519-528.
Webster C.R. & Lorimer C.G., 2005. Minimum opening sizes for canopy recruitment of midtolerant tree species: a retrospective approach. Ecol. Appl., 15(4), 1245-1262.
Pour citer cet article
A propos de : Yao Sadaiou Sabas Barima
Université Libre de Bruxelles (ULB). Service d'Écologie du Paysage et Systèmes de Production végétale. CP 169. Avenue Franklin Roosevelt, 50. B-1050 Bruxelles (Belgique). E-mail : ybarima@ulb.ac.be
A propos de : Nicolas Barbier
Université Libre de Bruxelles (ULB). Laboratoire de Complexité et Dynamique des Systèmes tropicaux. CP 169. Avenue Franklin Roosevelt, 50. B-1050 Bruxelles (Belgique).
A propos de : Brahima Ouattara
Université de Cocody-Abidjan. Laboratoire de Botanique. 22 BP 582. CI-Abidjan 22 (Côte d'Ivoire).
A propos de : Jan Bogaert
Université Libre de Bruxelles (ULB). Service d'Écologie du Paysage et Systèmes de Production végétale. CP 169. Avenue Franklin Roosevelt, 50. B-1050 Bruxelles (Belgique).