


- Accueil
- Volume 14 (2010)
- numéro spécial 2
- Fermentative hydrogen production by Clostridium butyricum CWBI1009 and Citrobacter freundii CWBI952 in pure and mixed cultures


Visualisation(s): 5579 (28 ULiège)
Téléchargement(s): 0 (0 ULiège)

Fermentative hydrogen production by Clostridium butyricum CWBI1009 and Citrobacter freundii CWBI952 in pure and mixed cultures

Résumé
Production d’hydrogène par Clostridium butyricum CWBI1009 et Citrobacter freundii CWBI952 en cultures pures et mixtes. Cet article étudie la production de biohydrogène par deux souches mésophiles, une anaérobie stricte (Clostridium butyricum CWBI1009) et une anaérobie facultative (Citrobacter freundii CWBI952). Ces souches ont été testées en cultures batch pures et mixtes sur cinq substrats différents. Les rendements de conversion des cultures pures de C. freundii s’étalent de 0.09 molH2∙molhexose-1 (sur saccharose) à 0.24 molH2∙molhexose-1 (sur glucose). Des rendements plus élevés sont atteints par Cl. butyricum, de 0.44 molH2∙molhexose-1 (sur saccharose) à 0.69 molH2∙molhexose-1 (sur lactose). Cette dernière souche dégrade l’amidon, tandis que C. freundii ne le consomme pas. Cependant, cette dernière assimile les autres substrats plus rapidement et produit de l’hydrogène plus tôt. Cet avantage a été utilisé pour promouvoir les conditions de croissance de Cl. butyricum en co-culture avec C. freundii, étant donné que Cl. butyricum est extrêmement sensible aux traces d’oxygène dans le milieu. Ceci permettrait d’éviter l’ajout d’agents réducteurs couteux dans le milieu de culture (tels que la L-cystéine) dans la mesure où C. freundii consomme l’oxygène résiduel. Ce type de co-culture a été testé sur glucose et sur amidon. Elles ont eu pour conséquence de diminuer les rendements de conversion sur glucose (de 0.53 molH2∙molhexose-1 pour les cultures pures de Cl. butyricum à 0.38 molH2∙molhexose-1 pour les cultures mixtes), tandis qu’ils ont légèrement augmenté sur amidon (passant respectivement de 0.69 and 0.73 molH2∙molhexose-1). Après 48 h de fermentation, les analyses métaboliques, confirmées par des observations microbiologiques, ont montré que le nombre de cellules de C. freundii ont drastiquement diminué ou que cette dernière souche a été très fortement inhibée par le développement de Cl. butyricum.
Abstract
This paper investigates the biohydrogen production by two mesophilic strains, a strict anaerobe (Clostridium butyricum CWBI1009) and a facultative anaerobe (Citrobacter freundii CWBI952). They were cultured in pure and mixed cultures in serum bottles with five different carbon sources. The hydrogen yields of pure C. freundii cultures ranged from 0.09 molH2∙molhexose-1 (with sucrose) to 0.24 molH2∙molhexose-1 (with glucose). Higher yields were obtained by the pure cultures of Cl. butyricum ranging from 0.44 molH2∙molhexose-1 (with sucrose) to 0.69 molH2∙molhexose-1 (with lactose). This strain also fermented starch whereas C. freundii did not. However, it consumed the other substrates faster and produced hydrogen earlier than Cl. butyricum. This ability has been used to promote the growth conditions of Cl. butyricum in co-culture with C. freundii, since Cl. butyricum is extremely sensitive to the presence of oxygen which strongly inhibits H2 production. This approach could avoid the addition of any expensive reducing agents in the culture media such as L-cysteine since C. freundii consumes the residual oxygen. Thereafter, co-cultures with glucose and starch were investigated: hydrogen yields decreased from 0.53 molH2∙molhexose-1 for pure Cl. butyricum cultures to 0.38 molH2∙molhexose-1 for mixed culture with glucose but slightly increased with starch (respectively 0.69 and 0.73 molH2∙molhexose-1). After 48 h of fermentation, metabolites analysis confirmed with microbial observation, revealed that the cell concentration of C. freundii dramatically decreased or was strongly inhibited by the development of Cl. butyricum.
Table des matières
1. Introduction
1Hydrogen (H2), whether burned or used directly in a fuel cell, is a very promising clean energy vector for the decrease of our environmental impact since its utilization generates only water vapor. Nevertheless, H2 is still mainly produced by steam reforming of methane, a process releasing large amount of fossil CO2 in the atmosphere. In the last few years, there has been an increasing interest to find new H2 production processes with almost no carbon emission (Balat, 2009; Holladay et al., 2009; Moriarty et al., 2009). One of the most promising and investigated prospects is the biological production of hydrogen through the degradation of a large spectrum of carbon sources by anaerobic bacteria in a process called “dark fermentation” (Das et al., 2001; Levin et al., 2004; Nath et al., 2004a; Das, 2009; Hallenbeck, 2009).
2The best described mesophilic strains are, on the one hand, strict anaerobic bacteria from the genus Clostridium that have the potential to reach high experimental hydrogen yields (about two moles of hydrogen per mole of hexose consumed). And on the other hand, facultative anaerobes such as Enterobacteriaceae that present lower experimental yields (~ 1 molH2.molhexose) but can achieve higher production rates (Hallenbeck et al., 2002; Hawkes et al., 2002; Kotay et al., 2008). The main purpose to enhance fermentative hydrogen production is to improve hydrogen yields for an efficient energy recovery from the substrate. The two species investigated in this work, Clostridium butyricum CWBI1009 (Masset et al., 2010) and Citrobacter freundii CWBI952 (Hamilton et al., 2010), have a maximum theoretical hydrogen yield of 4 and 2 molH2∙molhexose-1 respectively depending on the metabolic pathway followed for the fermentation of the carbon source (Nandi et al., 1998; Nath et al., 2004b; Kraemer et al., 2007; Oh et al., 2008a).
3Clostridia are however extremely sensitive to the presence of oxygen which strongly inhibits H2 evolving enzymes (Heinekey, 2009). This can be avoided with the addition of an expensive reducing agent such as L-cysteine. However, the use of such an agent is not suitable for a large-scale cost effective biohydrogen production process (Das et al., 2008; Yuan et al., 2008).
4Hydrogen may evolve through the fermentation processes of simple carbohydrates such as glucose, sucrose, lactose and maltose or more complex ones such as starch or even cellulose (Ueno et al., 1995; Davila-Vazquez et al., 2008; Magnusson et al., 2008). Only a few studies have investigated the hydrogen production with these different substrates on pure cultures in comparison with co-cultures (Yokoi et al., 1995; Nath et al., 2006; Chen et al., 2008; Pan et al., 2008). This is why this work compares the hydrogen and major metabolites production (i.e. acetate, butyrate, formate, lactate, ethanol and succinate) in pure C. freundii and Cl. butyricum cultures with five different substrates. These experiments were carried out in serum bottles batch cultures based on the biochemical hydrogen potential (BHP) test procedure described by Lin (Lin et al., 2007).
5Furthermore, a co-culture of C. freundii and Cl. butyricum in the same BHP culture may enhance hydrogen production. Such a culture would not require the addition of any reducing agents since C. freundii consumes oxygen and provides the anaerobic conditions required for Cl. butyricum growth. This has already been shown in a mixed culture of Enterobacter sp. and Cl. butyricum growing on starch (Yokoi et al., 1998; 2002). However, little is known about this consortium on other substrates. In this work, co-cultures were monitored on glucose and also on starch for comparison. They are discussed in comparison with the results found in the literature.
2. Materials and methods
2.1. Microorganisms conservation and preculture media
6The two species cultured in this work, i.e. Citrobacter freundii CWBI952 and Clostridium butyricum CWBI1009 were previously isolated and characterized in our laboratory (Hamilton et al., 2010; Masset et al., 2010). For the conservation of the strains 1 ml of a previous 25 ml culture, hermetically stoppered and incubated at 30°C, was transferred weekly to a fresh tube filled with sterile MD medium. The MD medium contained per liter of deionized water: glucose monohydrate (5 g), casein peptone (5 g), yeast extract (0.5 g), KH2PO4 (2 g), MgSO4.7H2O (0.5 g) and L-cysteine hydrochloride (0.5 g). All the chemicals used were of analytical or extra pure quality and were supplied by Merck, UCB and Sigma. Casein peptone and yeast extract were supplied by Organotechnie (La Courneuve, France).
2.2. Experimental procedures and culture media
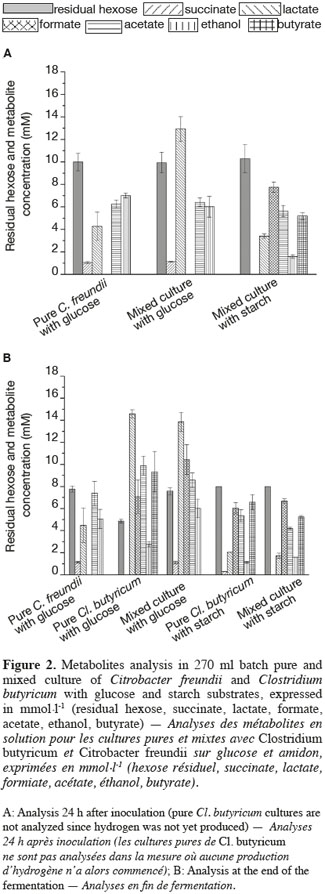
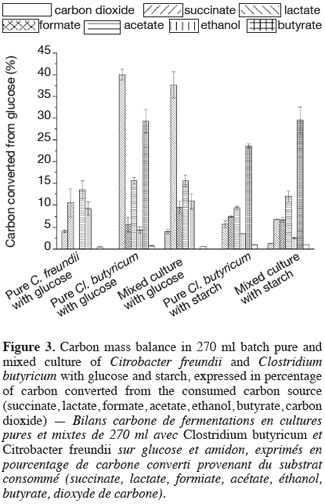
7The BHP (Biochemical Hydrogen Potential) tests were carried out in 270 ml sterile glass serum bottles filled with 200 ml of MD medium and different carbon sources in order to determine the hydrogen production potential of the strains. The sterile carbon source (i.e. glucose monohydrate, maltose, sucrose, lactose or starch in solution in deionized water) were prepared and added separately at a final concentration of 4.3 gCOD∙l-1 to prevent Maillard reactions between carbohydrates and amino acids. The same approach was followed with L-cysteine hydrochloride to a final concentration of 0.5 g∙l-1. The effective final substrate concentration was measured by HPLC. Differences between the initial concentrations of the carbon sources ranged from 3.9 to 4.8 gCOD∙l-1 due to hydration of the compounds. After the preparation of the MD medium and adjustment of the pH to 7.3 the bottles were sterilized. The carbon source and L-cysteine solution were then added before inoculation of the medium with 5 ml of a fresh preculture tube (incubated for 48 h) for the pure strains cultures and with 3 ml of each strain for the mixed cultures. The bottles were then capped with a butyl stopper as described by Lin (Lin et al., 2007) and flushed with nitrogen gas to remove the remaining oxygen in the bottle gas phase. The bottles were then incubated at 30°C ± 0.5°C with an orbital stirring at 150 rpm. Each BHP test was conducted in triplicate for the standard deviation estimations.
2.3. Analytical methods
8The cell concentration of C. freundii was determined by consecutive dilutions in peptoned water. A 100 µl sample of the three final dilutions was spread on PCA Petri dishes before incubation at 30°C for 18 to 24 h. This method was also used to confirm the absence of other microorganisms than C. freundii. The PCA (Plate Count Agar) medium contained per liter of deionized water: glucose monohydrate (1 g), casein peptone (5 g), yeast extract (2.5 g), agar (15 g). The cell concentration of Cl. butyricum or of the mixed culture were determined by microscopic observations on a Bürker counting chamber (mean of 10 counts) after dilution and cell settlement in a 0.4% (final concentration) fresh formaldehyde solution. The absence of aerobic contaminants was verified by spreading 100 µl of the culture on PCA medium plates and incubating for 48 h at 30°C. The absence of bacterial growth confirmed the absence of aerobic contaminants.
9The volume of biogas produced was measured with a sterile syringe and needle introduced through the previously sterilized septum. The proportion of hydrogen gas was determined using a gas chromatograph (GC) (Hewlett-Packard 5890 Series II) fitted with a thermal conductivity detector (TCD) and a 30 m x 0.32 mm GAS PRO GSC capillary column (Altech) in series with a 20 m x 0.25 mm CarboPLOT P7 column (Chrompak). The temperatures of the injection, TCD chambers and the oven were maintained at 90°, 110° and 55°C respectively. Nitrogen was used as the carrier gas in the column at a flow rate of 20 ml∙min-1.
10The liquid samples harvested during or at the end of the fermentations (ca. from 48 to 96 h) were centrifuged at 13,000 g for 10 min. The obtained supernatants were then filtered through a 0.2 µm cellulose acetate membrane (Midisart Sartorius) and analyzed by HPLC. The HPLC analysis was carried out using an Agilent 1100 series (HP Chemstation software) with a Supelcogel C-610H column preceded by a Supelguard H pre-column (oven temperature 40°C), 0.1% H3PO4 (in milliQ water) as the isocratic mobile phase at a flow rate of 0.5 ml∙min-1 and a differential refraction index detector (RID, heated at 35°C). The method lasted for 35 min at a maximum pressure of 60 bars. The concentrations of the compounds in the samples are determined in comparison with standard samples analyzed for the establishment of calibration curves. The data for the glucose (or any other carbon source) and metabolite concentrations were used to calculate the mass balance (MB) of the glucose conversion using the equation:

11where Ni is the number of carbon atoms in a molecule of metabolite i; ∆Ci is the concentration of metabolite i effectively produced (i.e. the difference between the final and initial concentrations); NG is the number of carbon atoms in the substrate molecule (i.e. 6) and ∆CG is the concentration of the glucose consumed (Eq. (1)).
3. Results and discussion
3.1. Hydrogen production by Citrobacter freundii CWBI952 and Clostridium butyricum CWBI1009 using five different substrates
12The investigation of hydrogen and metabolites production with five different substrates (glucose, maltose, sucrose, lactose and starch) was first carried out with pure cultures of Citrobacter freundii CWBI952 and Clostridium butyricum CWBI1009 in 270 ml batch serum bottles. Cell concentrations in the serum bottles at the beginning of the fermentation were of 4.3∙10+07 and 1.2∙10+07 CFU∙ml-1 respectively. The hydrogen production performances were compared in terms of hydrogen yields allowing a firm basis for a comparison of the two strains. The cumulative volumes of hydrogen produced and the hydrogen yields are reported in table 1 along with the substrate conversion ratios and the final cell concentrations (48 h after inoculation).
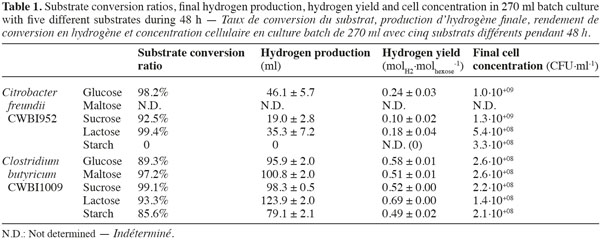
13These results clearly show that better performances were obtained with Cl. butyricum compared to C. freundii which produced hydrogen less efficiently. With starch C. freundii produced no hydrogen. This had already been reported in the literature and is due to its lack of functional amylases (Kotay et al., 2009). Although no hydrogen was produced by C. freundii, biomass increased to 3.3∙10+08 CFU∙ml-1 probably due to the consumption of other less accessible carbon sources in the media, i.e. casein peptone and yeast extract. By contrast Cl. butyricum is able to produce hydrogen by degrading starch.
14For every substrate investigated, higher hydrogen yields were obtained with Cl. butyricum. In addition, hydrogen was more abundant in the biogas produced by Cl. butyricum compared to C. freundii (on average 70% vs 62% respectively, data not reported here).
15The differences in the performances of the two strains are due to the different metabolic pathways followed (Ying et al., 2004; Oh et al., 2008b). Enterobacteriaceae produce hydrogen via the decomposition of formate up to a maximum yield of 2 molH2∙molhexose-1. Whereas Clostridia follows the mixed acid fermentation and provides a maximum yield that is twice as high when producing acetate and butyrate. However, the yields obtained in this investigation, ranging from 0.1 to 0.24 and 0.49 to 0.69 molH2∙molhexose-1 for C. freundii and Cl. butyricum respectively, are lower than those published in several studies [(0.88 molH2∙molhexose-1 for C. freundii (Hamilton et al., 2010) and 1.69 molH2∙molhexose-1 for Cl. butyricum (Masset et al., 2010)]. This difference can be explained by the absence of pH regulation in the serum bottle experiments. During the fermentation of the carbon sources, acid metabolites were produced leading to a rapid decrease in pH (set at 7.3 at the beginning). Since no buffer such as HPO42-/ H2PO4- was used in the media the pH of the cultures rapidly dropped to levels lower than the optimal pH for hydrogen production (5.9 and 5.2 for C. freundii and Cl. butyricum respectively) thereby reducing the final yield reached. In addition the final pH levels were much lower (ranging from 4.13 to 4.67) inducing a strong inhibitory effect, not only on the hydrogen production but also on substrate consumption.
3.2. Metabolites analysis for C. freundii and Cl. butyricum with different substrates
16The main soluble metabolites (ethanol, lactate, acetate, succinate and formate for C. freundii and butyrate, lactate, acetate, formate and ethanol for Cl. butyricum) were analyzed by HPLC at the end of the culture (48 h). The carbon mass balance for these metabolites is indicated in the figure 1 in terms of the percentage of substrate’s carbon content converted. Due to the difference in the metabolic pathways involved, butyrate is only produced by Cl. butyricum and succinate only by C. freundii. In addition more of the carbon source is converted to ethanol by C. freundii (ten times more than with Cl. butyricum).
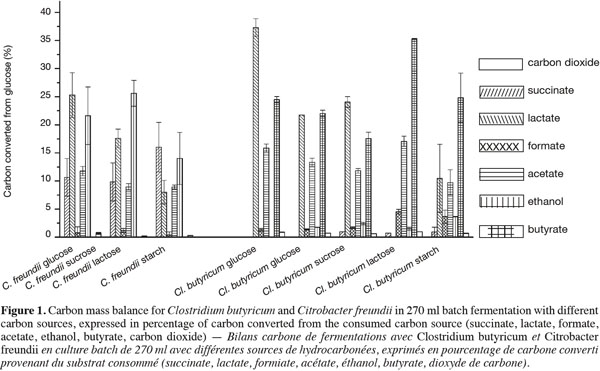
17With the C. freundii cultures only small amounts of formate were detected indicating that only a little more hydrogen, enhancing the yields by about 20%, could have been produced through the activity of the Formate Hydrogen Lyase (FHL) enzymatic complex decomposing formate in hydrogen (Kim et al., 2008). Other metabolites, mainly ethanol and lactate accounting for about 20% of the glucose converted, are of less interest in this context since they do not lead to the production of hydrogen. When the cultures were maintained in optimal conditions, these metabolites were still produced in similar amounts. This indicates that very little or no adaptation in C. freundii’s metabolic pathway occurs in response to these conditions (Hamilton et al., 2010).
18With Clostridia’s metabolic pathway, only acetate and butyrate lead to the coproduction of hydrogen, accounting for 15.9% and 24.5% respectively of the glucose converted. In a previous work, regulated batch cultures at the optimal pH for hydrogen production were carried out with Cl. butyricum and these two metabolites accounted for 12.7% and 40.3% respectively with a yield of 1.69 molH2∙molhexose-1 (Masset et al., 2010). The lower conversion rate of carbohydrate into acetate and butyrate shown here explains the low hydrogen yields obtained in the serum bottles cultures. Therefore, it can be assumed that Cl. butyricum adapts its metabolism to produce more favorable metabolites and hydrogen when growing under optimal pH conditions. The synthesis of lactate is not accompanied by hydrogen production. In this study, no lactate was produced from lactose by Cl. butyricum. This observation may be linked to the higher hydrogen yields reached compared to other substrates (Table 1) and to the higher conversion rate of the carbon source into butyrate (35.3% with lactose and 24.5% with glucose). By contrast, a considerable amount of lactate was produced from the other carbohydrates (e.g. 37.3% of the initial glucose). Cl. butyricum also synthesizes formate accounting for 2 to 3% of the substrate. This metabolite could be used in a co-culture of the two species to potentially produce more hydrogen via C. freundii’s metabolism. Yokoi has already investigated this topic, reporting that such a mixed culture produced hydrogen efficiently and without the need to add an expensive reducing agent (Yokoi et al., 1998).
3.3. Batch fermentation with co-culture of C. freundii and Cl. butyricum on glucose and starch
19Mixed cultures of Cl. butyricum and C. freundii were cultured in 270 ml serum bottles with glucose or starch. Pure cultures were carried out in the same conditions for comparison. L-cysteine was added in each culture according to the media description in materials and methods. The results of these experiments are reported in table 2 indicating the hydrogen production measured after 24 h of culture and at the end of the fermentation (96 h on glucose and 48 h on starch) and hydrogen yields.

20The results of these experiments with starch are in accordance with the results obtained by Yokoi: a sustainable hydrogen production, in terms of hydrogen production and yield, is possible with starch (Yokoi et al., 2002). In our cultures C. freundii seemed to survive and, as reported by Yokoi, was subsequently able to re-establish viable growth condition for Cl. butyricum in case of oxygen contamination in the culture media. The results presented in the table 2 suggest that C. freundii establishes anaerobic conditions and allows production of hydrogen to begin earlier than in a pure Cl. butyricum culture (54 ml were already produced 24 h after inoculation).
21To our knowledge such a sustainable consortium with a simple sugar such as glucose has not yet been studied. During the first 24 h of fermentation no hydrogen was produced by Cl. butyricum while C. freundii had already provided more than 95% of its total hydrogen production. The sustainable influence of C. freundii in the mixed culture with glucose allowed the production of hydrogen to begin earlier than with the pure Cl. butyricum culture. However, the use of a mixed culture in batch fermentation decreased the final hydrogen yield from 0.53 to 0.39 molH2∙molhexose-1 by comparison between the mixed culture and the pure Cl. butyricum culture. Tests on PCA media, carried out 24 h after inoculation, did not show any aerobic bacterial development indicating that C. freundii did not survive or that the cell concentration had dramatically decreased. This suggests that the strain enhanced the initiation of hydrogen production, but was then rapidly overgrown or strongly inhibited by Cl. butyricum.
22This is confirmed by the results presented in figures 2 and 3 depicting the analysis of metabolites and carbon mass balance. They show that lactate and butyrate are the main metabolites from the degradation of glucose and starch. These two metabolites are commonly found in Cl. butyricum culture supernatants. However, the presence of succinate indicates that C. freundii grew at the beginning of the fermentation.
23The data obtained on starch degradation confirm that the metabolites (high butyrate and low ethanol concentrations) seemed to correspond to the fermentation performed by Cl. butyricum and that C. freundii was strongly inhibited under these conditions.
24The production of succinate and the high amount of ethanol detected in the supernatants of the glucose media indicates that C. freundii was active at the beginning of the fermentation and grew faster than Cl. butyricum consuming part of the glucose. This would explain the decrease in the final hydrogen yields (see table 2). As C. freundii was not found in the media at the end of the fermentation, it had by then been overgrown by Cl. butyricum. Inhibition of C. freundii growth may be a limiting factor if a sudden infiltration of oxygen occurs in the absence of a reducing agent, since the strictly anaerobic strain Cl. butyricum may then start to sporulate.
4. Conclusion
25Our work highlights the fact that mixed cultures of Citrobacter freundii CWBI952 and Clostridium butyricum CWBI1009 can efficiently maintain the production of hydrogen at acceptable yields compared to pure cultures of Cl. butyricum. Moreover, it could consume efficiently many different carbon sources since Cl. butyricum was able to degrade simple carbohydrates or even starch. However the survival of C. freundii in competition with Cl. butyricum was compromised since it was overgrown especially with the glucose substrate. Further work will be done to find other facultative aerobic strains which are able to enhance anaerobic conditions in the culture media without being overgrown by Cl. butyricum and without decreasing the hydrogen yield.
26Acknowledgements
27L. Beckers and S. Hiligsmann both equally contributed to the paper. L. Beckers is grateful to the FRS-FNRS (Fonds de la Communauté française de Belgique pour la Recherche Scientifique) for supporting his work and researches. C. Hamilton is to be thanked for his guidance through the redaction of this paper. J. Masset is a recipient of a FRIA fellowship (fonds pour la Formation à la Recherche dans l’Industrie et l’Agriculture). This work was also supported by an ARC project (Action de Recherches Concertées, ARC-07/12-04) and the Walloon Region. The publication of this article has been allowed by the organization of the second thematic day organized by the GEPROC doctoral thematic school.
Bibliographie
Balat M., 2009. Possible methods for hydrogen production. Energy Sources Part A Recovery Util. Environ. Effects, 31(1), 39-50.
Chen S.D. et al., 2008. Batch and continuous biohydrogen production from starch hydrolysate by Clostridium species. Int. J. Hydrogen Energy, 33(7), 1803-1812.
Das D., 2009. Advances in biohydrogen production processes: an approach towards commercialization. Int. J. Hydrogen Energy, 34(17), 7349-7357.
Das D. & Veziroglu T.N., 2001. Hydrogen production by biological processes: a survey of literature. Int. J. Hydrogen Energy, 26(1), 13-28.
Das D. & Veziroglu T. N., 2008. Advances in biological hydrogen production processes. Int. J. Hydrogen Energy, 33(21), 6046-6057.
Davila-Vazquez G. et al., 2008. Fermentative biohydrogen production: trends and perspectives. Rev. Environ. Sci. Biotechnol., 7(1), 27-45.
Hallenbeck P.C., 2009. Fermentative hydrogen production: principles, progress, and prognosis. Int. J. Hydrogen Energy, 34(17), 7379-7389.
Hallenbeck P.C. & Benemann J.R., 2002. Biological hydrogen production; fundamentals and limiting processes. Int. J. Hydrogen Energy, 27(11-12), 1185-1193.
Hamilton C. et al., 2010. Optimization of culture conditions for biological hydrogen production by Citrobacter freundii cwbi952 in batch, sequenced-batch and semicontinuous operating mode. Int. J. Hydrogen Energy, 35(3), 1089-1098.
Hawkes F.R., Dinsdale R., Hawkes D.L. & Hussy I., 2002. Sustainable fermentative hydrogen production: challenges for process optimisation. Int. J. Hydrogen Energy, 27(11-12), 1339-1347.
Heinekey D.M., 2009. Hydrogenase enzymes: recent structural studies and active site models. J. Organomet. Chem., 694(17), 2671-2680.
Holladay J.D., Hu J., King D.L. & Wang Y., 2009. An overview of hydrogen production technologies. Catal. Today, 139(4), 244-260.
Kim S. et al., 2008. Various hydrogenases and formate-dependent hydrogen production in Citrobacter amalonaticus y19. Int. J. Hydrogen Energy, 33(5), 1509-1515.
Kotay S.M. & Das D., 2008. Biohydrogen as a renewable energy resource - prospects and potentials. Int. J. Hydrogen Energy, 33(1), 258-263.
Kotay S.M. & Das D., 2009. Novel dark fermentation involving bioaugmentation with constructed bacterial consortium for enhanced biohydrogen production from pretreated sewage sludge. Int. J. Hydrogen Energy, 34(17), 7489-7496.
Kraemer J.T. & Bagley D.M., 2007. Improving the yield from fermentative hydrogen production. Biotechnol. Lett., 29(5), 685-695.
Levin D.B., Pitt L. & Love M., 2004. Biohydrogen production: prospects and limitations to practical application. Int. J. Hydrogen Energy, 29(2), 173-185.
Lin P.Y. et al., 2007. Biological hydrogen production of the genus Clostridium: metabolic study and mathematical model simulation. Int. J. Hydrogen Energy, 32, 1728-1735.
Magnusson L. et al., 2008. Direct hydrogen production from cellulosic waste materials with a single-step dark fermentation process. Int. J. Hydrogen Energy, 33(20), 5398-5403.
Masset J. et al., 2010. Effect of ph on glucose and starch fermentation in batch and sequenced-batch mode with a recently isolated strain of hydrogen-producing Clostridium butyricum cwbi1009. Int. J. Hydrogen Energy, 35(8), 3371-3378.
Moriarty P. & Honnery D., 2009. Hydrogen's role in an uncertain energy future. Int. J. Hydrogen Energy, 34(1), 31-39.
Nandi R. & Sengupta S., 1998. Microbial production of hydrogen: an overview. Crit. Rev. Microbiol., 24(1), 61-84.
Nath K. & Das D., 2004a. Biohydrogen production as a potential energy resource - present state-of-art. J. Sci. Ind. Res., 63(9), 729-738.
Nath K. & Das D., 2004b. Improvement of fermentative hydrogen production: various approaches. Appl. Microbiol. Biotechnol., 65(5), 520-529.
Nath K., Kumar A. & Das D., 2006. Effect of some environmental parameters on fermentative hydrogen production by enterobacter cloacae dm11. Can. J. Microbiol., 52(6), 525-532.
Oh Y.K. et al., 2008a. Metabolic-flux analysis of hydrogen production pathway in Citrobacter amalonaticus y19. Int. J. Hydrogen Energy, 33(5), 1471-1482.
Oh Y.K. et al., 2008b. Carbon and energy balances of glucose fermentation with hydrogen-producing bacterium Citrobacter amalonaticus y19. J. Microbiol. Biotechnol., 18(3), 532-538.
Pan C.M., Fan Y.T., Zhao P. & Hou H.W., 2008. Fermentative hydrogen production by the newly isolated Clostridium beijerinckii fanp3. Int. J. Hydrogen Energy, 33(20), 5383-5391.
Ueno Y. et al., 1995. Biological production of hydrogen from cellulose by natural anaerobic microflora. J. Ferment. Bioeng., 79(4), 395-397.
Ying Z. & Yang S.T., 2004. Effect of ph on metabolic pathway shift in fermentation of xylose by Clostridium tyrobutyricum. J. Biotechnol., 110(2), 143-157.
Yokoi H. et al., 1995. Characteristics of hydrogen production by aciduric enterobacter aerogenes strain ho-39. J. Ferment. Bioeng., 80(6), 571-574.
Yokoi H. et al., 1998. H2 production from starch by a mixed culture of Clostridium butyricum and enterobacter aerogenes. Biotechnol. Lett., 20(2), 143-147.
Yokoi H., Maki R., Hirose J. & Hayashi S., 2002. Microbial production of hydrogen from starch-manufacturing wastes. Biomass Bioenergy, 22(5), 389-395.
Yuan Z.L., Yang H.J., Zhi X.H. & Shen J.Q., 2008. Enhancement effect of l-cysteine on dark fermentative hydrogen production. Int. J. Hydrogen Energy, 33(22), 6535-6540.
Pour citer cet article
A propos de : Laurent Beckers
Univ. Liege (ULg). Walloon Centre of Industrial Biology (WCIB). Boulevard du Rectorat, 29/40-P.70. B-4000 Liege (Belgium). E-mail: lbeckers@ulg.ac.be
A propos de : Serge Hiligsmann
Univ. Liege (ULg). Walloon Centre of Industrial Biology (WCIB). Boulevard du Rectorat, 29/40-P.70. B-4000 Liege (Belgium).
A propos de : Christopher Hamilton
Univ. Liege (ULg). Walloon Centre of Industrial Biology (WCIB). Boulevard du Rectorat, 29/40-P.70. B-4000 Liege (Belgium).
A propos de : Julien Masset
Univ. Liege (ULg). Walloon Centre of Industrial Biology (WCIB). Boulevard du Rectorat, 29/40-P.70. B-4000 Liege (Belgium).
A propos de : Philippe Thonart
Univ. Liege (ULg). Walloon Centre of Industrial Biology (WCIB). Boulevard du Rectorat, 29/40-P.70. B-4000 Liege (Belgium).