


- Accueil
- volume 12 (2008)
- Rôle de l'intensité lumineuse sur les capacités parasitaires d'Eupelmus orientalis Crawford et d'Eupelmus vuilleti Crawford, parasitoïdes des Bruchidae ravageurs de graines de niébé (Vigna unguiculata Walp.)


Visualisation(s): 1498 (118 ULiège)
Téléchargement(s): 42 (16 ULiège)

Rôle de l'intensité lumineuse sur les capacités parasitaires d'Eupelmus orientalis Crawford et d'Eupelmus vuilleti Crawford, parasitoïdes des Bruchidae ravageurs de graines de niébé (Vigna unguiculata Walp.)

Notes de la rédaction
Reçu le 9 juin 2006, accepté le 4 septembre 2007
Résumé
En Afrique tropicale, Callosobruchus maculatus Fabricius est un ravageur du niébé. Après la récolte, le temps de conservation des graines est limité par la disparition des stocks d'un des parasitoïdes (Eupelmus orientalis Crawford) de C. maculatus. Au cours de cette étude, nous avons utilisé un modèle expérimental qui simule les greniers traditionnels africains en tenant compte du manque d'étanchéité et de la variabilité de l'intensité lumineuse interne des différents types de greniers. Trois types de cages ont été utilisés correspondant à trois niveaux d'intensité lumineuse : 380 lux, 3,5 lux et 0,2 lux. Quatre petits orifices ont été aménagés sur les parois de chaque cage, chacun étant muni d'un piège pour attraper les insectes qui s'évadent. A l'intérieur de chaque cage, nous avons placé 20 nymphes de E. orientalis, 20 de Eupelmus vuilleti Crawford et 240 larves de C. maculatus. Au stade adulte, les insectes ont le choix entre s'évader ou bien rester dans la cage et éventuellement parasiter les hôtes disponibles. Les résultats montrent que les femelles E. orientalis nées dans les cages parasitent peu les hôtes disponibles et s'évadent majoritairement par rapport à celles de E. vuilleti qui restent dans les cages. Dans les enceintes de 380 lux, 19 % des femelles E. orientalis s'évadent contre 6 % pour E. vuilleti. Ce pourcentage d'évasion augmente avec l'obscurcissement de l'enceinte. Ainsi, 62 % des femelles E. orientalis s'évadent des cages de 3,5 lux et 67 % des cages de 0,2 lux, alors qu'aucune femelle E. vuilleti ne s'échappe des cages de 3,5 lux et seulement 11 % s'évadent des cages de 0,2 lux. La plupart des individus E. orientalis s'évadent dès l'émergence, cela explique donc leur disparition des stocks. Ce comportement d'évasion constitue un facteur limitant pour l'utilisation de ce parasitoïde comme auxiliaire de lutte contre les bruches.
Abstract
Role of light intensity on parasitic capacities of Eupelmus orientalis Crawford and Eupelmus vuilleti Crawford, parasitoids of Bruchidae pests of cowpea (Vigna unguiculata Walp.) seeds. In tropical Africa, Callosobruchus maculatus Fabricius is a cowpea pest. After the harvest, the disappearance of one of C. maculatus parasitoids (Eupelmus orientalis Crawford) limits the duration of stocks conservation. In this study, we used an experimental model imitating the traditional african granaries taking into account the lack of airtightness and the variability of the internal luminous intensity of the different types of granaries. Three kinds of cages were used: cages with an internal luminous intensity of 380 lux, cages with an internal luminous intensity of 3.5 lux, and some others with an internal luminous intensity of 0.2 lux. Four little holes are pierced on the sides of each cage. Each hole was provided with a trap to catch the insects that escaped. Inside each cage, 20 E. orientalis and Eupelmus vuilleti Crawford nymphs were placed. In adult stage, the insects faced a choice of either to escape, or to stay in the cage and then parasitize available hosts. The trapped individuals as well as those remaining in the cage are sorted out by species. Available hosts in the cage which had parasites are also sorted out. The results show that the E. orientalis females born in the cage hardly parasitize available hosts and escape in majority compared to E. vuilleti which stay in the cages. In 380 lux cages, 19% of E. orientalis females escape compared to 6% of E. vuilleti. This percentage of evasion increases with the darkning of the enclosure. Thus, 62% of E. vuilleti females escape from the cages of 3.5 lux and only 11% escape from the cages of 0.2 lux. The majority of E. orientalis individuals escape right from birth, which therefore explains their disappearance from storage. This escape behaviour constitutes a factor limiting the use of this parasitoid as a biological control agent against bruchids.
Table des matières
1. Introduction
1En milieu rural ouest africain, le niébé est stocké en gousses ou en graines dans les greniers traditionnels en banco (mélange d'argile et de paille) ou en paille (tiges de mil) après les récoltes. La période de stockage peut durer 9 à 10 mois selon l'importance de la production (Idi, 1994). Le principal problème des agriculteurs est la conservation des récoltes dont 80 à 100 % sont détruits par le développement de deux espèces de bruches (Bruchidius atrolineatus Pic et Callosobruchus maculatus Fabricius) 2 à 3 mois après le début du stockage.
2Dans certaines régions d'Afrique, notamment au Sahel, trois espèces de parasitoïdes larvaires (Dinarmus basalis Rondoni, Eupelmus vuilleti Crawford et Eupelmus orientalis Crawford) sont associées aux populations de bruches (Alzouma, 1987 ; Germain, 1988 ; Monge et al., 1991). Ces parasitoïdes sont en proportions variables selon les années, la seule constante au cours des ans étant que E. orientalis ne se maintient pas dans les systèmes de stockage. Bien qu'elle soit introduite avec des gousses contaminées par C. maculatus dans les champs, la population de ce parasitoïde régresse rapidement au profit de D. basalis et surtout de E. vuilleti qui finit par dominer (Monge et al., 1991).
3Des études menées par Lévêque et al. (1993) ont montré qu'il existe une compétition entre D. basalis et E. vuilleti. Cette compétition est responsable de la diminution des effectifs de D. basalis (Monge et al., 1995) et du maintien dans les stocks des populations de bruches à des taux indésirables pour les agriculteurs. La question était de savoir si la compétition entre E. orientalis et E. vuilleti se faisait aussi au détriment d'E. orientalis.
4D'autres travaux ont montré que la disparition d'E. orientalis des greniers ne pouvait être imputée ni à une réduction de ses potentialités reproductrices ni à un comportement agressif de E. vuilleti, puisque les femelles de cette espèce évitent les hôtes déjà parasités par E. orientalis (Ndoutoume, 1996). Que devient donc E. orientalis une fois entré dans les greniers après les récoltes ? Les matériaux de construction des greniers traditionnels (banco ou paille) n'en font pas des enceintes étanches. Les greniers en paille et, dans une moindre mesure, ceux en banco, ont des orifices par lesquels peuvent s'évader ou pénétrer les insectes. Dans ce type d'habitat mi-clos où se côtoient plusieurs espèces de parasitoïdes, une espèce généraliste comme E. orientalis, rencontrée en forêt, en savane et dans des cultures (Rasplus, 1988), trouve dans le grenier un habitat peu favorable compte tenu de ses exigences écologiques. Sachant que dans les champs E. orientalis est l'espèce la plus abondante du cortège parasitaire des bruches, nous avons émis l'hypothèse qu'elle pourrait éviter la compétition en espace clos en s'évadant des structures de stockage. C'est cette hypothèse que nous avons vérifiée lors de cette étude en conditions de laboratoire.
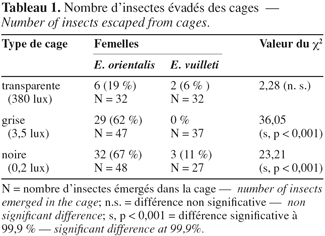
2. Matériel et méthodes
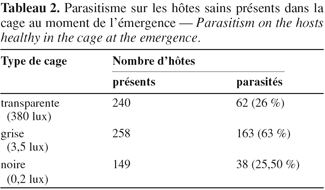
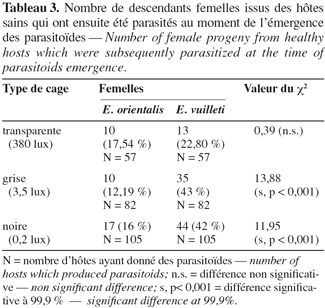
5Les insectes utilisés au cours de cette étude sont issus des élevages de masse de C. maculatus et de ses parasitoïdes Eupelmidae (E. vuilleti et E. orientalis) maintenus dans une pièce climatisée où les conditions abiotiques sont contrôlées [L.D. 12/12 (intensité lumineuse de 380 lux pendant la photophase) ; 32°/22°C ; 50 % : 80 % H.R.]. Les élevages des parasitoïdes ont été initiés à partir d'adultes émergés de graines de Vigna unguiculata contaminées par C. maculatus et collectées dans la région de Niamey (Niger). Sous les conditions abiotiques de la salle d'élevage, le stade larvaire L3 de C. maculatus s'observe en moyenne 11 à 15 jours après la ponte de l'œuf, le stade larvaire L4-prénymphe entre 15 et 18 jours, le stade nymphal entre 18 et 24 jours. L'émergence des adultes s'étale en moyenne entre 24 et 30 jours (Terrasse et al., 1986).
6Chez les parasitoïdes E. vuilleti et E. orientalis, le stade de l'hôte influe fortement sur l'activité reproductrice des femelles. Plus son développement est avancé, plus le nombre d'œufs pondus par jour croît et plus le sex-ratio des descendants est biaisé en faveur des filles (Terrasse et al., 1986 ; Doury et al., 1994). En présence d'hôtes du dernier stade larvaire et/ou de nymphes, les femelles de E. orientalis engendrent 65,67 % de filles et celles de E. vuilleti 63 % (Doury et al., 1994 ; Ndoutoume et al., 2000). Ces Eupelmidae se reproduisent par amphimixie mais aussi par parthénogenèse arrhénotoque et les mâles des deux espèces sont identiques. En raison de cette parthénogenèse, s'il y a absence de mâles, quelques femelles suffisent pour reconstituer, en une génération, une population à deux sexes en présence d'hôtes. C'est pour cela que l'impossibilité de différencier les mâles des deux espèces ne constitue pas un facteur qui biaise l'activité parasitaire et les comportements associés (rythmes d'activité, compétition, évasion, etc.) car seules les femelles parasitent.
2.1. Modèle expérimental
7En Afrique, l'utilisation de plusieurs types de matériaux (tiges de mil, sorgho, argile et autres) pour la construction des greniers fait que l'intensité de la lumière qui y pénètre varie selon le matériau utilisé. Compte tenu des différences de réponse à la lumière des femelles d'E. orientalis et d'E. vuilleti (Ndoutoume et al., 2006), il est probable que le comportement des parasitoïdes soit influencé par l'intensité lumineuse à l'intérieur des stocks. Dans les expériences, il a été tenu compte des différences d'intensité lumineuse à l'intérieur des cages représentant les greniers (Ndoutoume, 1996). Trois types de cages en plexiglas ont été utilisés :
8– 1 cage transparente : intensité lumineuse interne de 380 lux
9– 1 cage peinte en gris : intensité lumineuse interne de 3,5 lux
10– 1 cage peinte en noir : intensité lumineuse interne de 0,2 lux
11Quatre petits orifices ont été aménagés sur les parois de chaque cage. Chaque orifice était muni d'un piège constitué par un petit tube en plastique dans lequel tombent les insectes qui tentent de s'échapper des cages. Pour chaque type de cage, six répétitions ont été réalisées.
12Ces trois types de cages diffèrent uniquement par l'intensité de la lumière interne. Tous les autres paramètres (dimensions, nombre d'hôtes et de femelles de parasitoïdes) sont identiques. Les cages mesurent 27 cm (L) × 17,5 cm (l) × 17,5 cm (h). Dans chaque cage, on a placé 40 hôtes renfermant chacun une nymphe d'Eupelmus (parasités 15 jours auparavant par des femelles accouplées). Parmi les 40 hôtes, 20 contiennent des nymphes de E. orientalis et les 20 autres contiennent des nymphes de E. vuilleti. Ces 40 hôtes sont disséminés au milieu de 240 hôtes sains (larves de bruches de stade 3). L'ensemble a été divisé en 4 lots (10 hôtes parasités au milieu de 60 hôtes sains). Deux des lots renferment des hôtes parasités par E. orientalis et les deux autres ceux parasités par E. vuilleti.
13Pour chacune des espèces de parasitoïde, un lot était réservé à l'étude de la répartition de la ponte et l'autre au dénombrement de la descendance. Cet ensemble d'hôtes (sains et parasités) peut évoluer jusqu'à la nymphose et à l'émergence des adultes. L'âge des deux catégories d'hôtes a été choisi de sorte qu'à l'émergence des parasitoïdes adultes, les hôtes n'aient pas terminé leur développement. Les parasitoïdes adultes disposaient ainsi, à l'émergence, de différents stades d'hôtes (larves de stade 3-4 ou nymphe) susceptibles d'être parasités. Les femelles des parasitoïdes pourront :
14– rester dans l'enceinte et parasiter ou non les hôtes disponibles,
15– s'évader de l'enceinte (après avoir ou non parasité les hôtes) par un des quatre petits orifices aménagés sur les parois.
16L'expérience a duré 15 jours et a pris fin à l'émergence des adultes de bruches. A ce moment-là, les parasitoïdes restant dans l'enceinte ont moins de chance de rencontrer des hôtes susceptibles d'être parasités. Les cages ont été maintenues à 32 C°/22 °C, 50 %/80 % H.R., L. D. 12/12 durant toute la période d'observation.
2.2. Paramètres relevés
17Au cours des 15 jours d'expérience, les pièges ont été relevés 4 fois par jour (2 fois dans la matinée et 2 fois dans l'après-midi). Les individus piégés sont dénombrés par sexe et par espèce pour ce qui concerne les femelles. A l'arrêt de l'expérience, tous les individus restés dans l'enceinte ont été comptés par espèce. Les graines destinées à l'étude de la ponte par les femelles parasitoïdes nées dans l'enceinte expérimentale ont été ouvertes pour permettre le dénombrement des hôtes parasités. Les lots destinés à l'étude ont été isolés et conservés dans les mêmes conditions thermo- et photopériodiques que celles de l'expérience jusqu'à l'émergence de tous les insectes qui ont été comptabilisés par sexe et par espèce pour ce qui concerne les femelles. Le dénombrement des hôtes parasités au sein des lots d'hôtes sains offerts aux parasites a permis d'évaluer l'activité parasitaire des femelles nées dans l'enceinte. Compte tenu de l'impossibilité d'évaluer l'activité parasitaire de chaque espèce par l'examen de la ponte, les œufs des deux espèces étant identiques, la descendance observée a été le paramètre utilisé. Il ne peut émerger qu'un parasitoïde par hôte car ces Eupelmidae sont des ectoparasitoïdes solitaires. Le test du χ² a permis de comparer l'importance de la descendance de chacune des deux espèces, ce qui donne une idée du nombre de femelles qui se sont reproduites sans quitter l'enceinte et dont l'effectif dépendait du nombre de femelles s'étant échappées.
3. Résultats
3.1. Evasions
18Le tableau 1 donne la proportion d'individus qui se sont échappés de chaque type de cage simulant un grenier. Les femelles E. orientalis s'évadent en plus grand nombre que celles d'E. vuilleti. Dans les enceintes les plus claires (380 lux), 19 % des femelles E. orientalis émergées s'évadent contre 6 % pour E. vuilleti. Ce pourcentage augmente avec l'obscurcissement de l'enceinte. Ainsi, 62 % des femelles E. orientalis s'évadent des cages grises (3,5 lux) et 67 % des cages noires (0,2 lux), alors qu'aucune femelle E. vuilleti ne s'échappe des cages grises et seulement 11 % s'évadent des cages noires.
3.2. Parasitisme des hôtes disponibles et descendance
19Certains des hôtes disponibles dans les cages au moment de l'émergence des parasitoïdes ont été parasités. Cela signifie que les femelles émergées dans les greniers ont une activité de parasitisme quelle que soit l'intensité lumineuse (Tableau 2). La part prise par chaque espèce dans ce parasitisme n'est connue que par l'analyse des descendants engendrés puisqu'il n'est pas possible de faire la différence entre les œufs des deux espèces. Le tableau 3 montre que dans les enceintes claires (380 lux), 17,54 % de descendants sont des femelles E. orientalis avec 22,80 % de femelles E. vuilleti. Dans les cages grises (3,5 lux), ces valeurs sont respectivement de 12,19 % et 43 %. Dans les cages noires (0,2 lux), pour l'espèce E. orientalis, le pourcentage de descendants est de 16 % alors qu'il atteint 42 % pour les femelles de E. vuilleti. Quelle que soit l'intensité lumineuse, les pourcentages de descendants femelles E. orientalis sont toujours plus faibles que ceux d'E. vuilleti et l'écart augmente avec l'obscurcissement de l'enceinte. Cela voudrait dire que les femelles d'E. orientalis s'évadent majoritairement plutôt que de pondre et laisser des descendants dans les cages.
4. Discussion et conclusion
20Le comportement d'évasion des deux Eupelmidae a été étudié avec trois intensités lumineuses à l'intérieur des cages. Les orifices aménagés pour la sortie des parasitoïdes représentent des points lumineux que sont les ouvertures qu'on retrouve sur les parois des greniers africains. Ces modèles expérimentaux simulent les conditions qu'ils peuvent rencontrer dans leur milieu naturel. Dans ce type d'habitat mi-clos, l'invasion progressive des graines stockées par les Bruchidae assure la stabilité des ressources aux trois espèces de parasitoïdes larvaires D. basalis, E. vuilleti, E. orientalis qui se côtoient dans cet habitat (Caswell, 1974 ; Huignard et al., 1985 ; Jackai et al., 1986 ). Dans un tel environnement, les effectifs de femelles, la quantité et la qualité des hôtes (parasités ou non) induisent de fortes pressions compétitives intra- et inter- spécifiques (Rojas-Rousse et al., 1999). Malgré cela, des échantillonnages périodiques dans les greniers révèlent la présence permanente de l'espèce D. basalis alors que les interactions D. basalis–E. vuilleti sont toujours défavorables à D. basalis du fait du cleptoparasitisme pratiqué par E. vuilleti vis-à-vis des œufs, des larves néonates et du premier stade larvaire de l'espèce concurrente (Lévêque et al., 1993). La cohabitation entre D. basalis et E. vuilleti est possible du fait de leur occupation différente de l'habitat : D. basalis parasite de préférence les hôtes dispersés dans la profondeur d'un stock expérimental alors que E. vuilleti préfère les hôtes distribués en surface (Ndoutoume, 1996 ; Cortesero et al., 1997).
21La pression de sélection, exercée par la qualité et la quantité d'hôtes disponibles, amène les femelles E. vuilleti et E. orientalis à pousser le comportement de cleptoparasitisme jusqu'à l'hyperparasitisme du dernier stade larvaire (Rojas-Rousse et al., 1999 ; Rojas-Rousse et al., 2005). Le passage du parasitisme primaire vers l'hyperparasitisme facultatif, avec le cleptoparasitisme comme étape intermédiaire, s'est produit sans nécessité d'une adaptation spéciale pour pondre et se nourrir aux dépens d'un hôte secondaire. L'hyperparasitisme facultatif, en induisant un niveau trophique supplémentaire, conduit à des adultes miniaturisés du fait de la baisse pondérale de la quantité de nourriture disponible de l'hôte secondaire (Kalmès et al., 2002 ; Rojas-Rousse et al., 2005).
22La disparition à partir du deuxième mois de stockage de l'espèce E. orientalis ne peut pas être due à une issue de la compétition défavorable à E. orientalis. En effet, les femelles E. vuilleti émergent et sont actives plusieurs heures avant le signal lumineux alors que les femelles E. orientalis émergent exclusivement pendant la photophase et ont un rythme d'activité très dépendant de la lumière (Ndoutoume et al., 2006). Cette différence de sensibilité au signal photopériodique induit un décalage du démarrage de l'activité locomotrice au cours de la journée, ce qui permet de réduire les contacts entre les adultes des deux espèces (Ndoutoume et al., 2006). Des croisements entre deux souches d'origine différente du parasitoïde Leptopilina heterotoma (Eucoilidae) ont démontré que les différences de sensibilité au signal photopériodique avaient des bases génétiques, ce qui suggère une adaptation du comportement de recherche des hôtes dans les populations naturelles (Fleury et al., 1995).
23La sensibilité au signal photopériodique des deux espèces d'Eupelmidae conduit une partie des individus (mâles et femelles) nés dans les enceintes expérimentales, à les quitter quelle que soit l'intensité lumineuse interne. L'activité locomotrice de E. orientalis, très dépendante de la lumière, est confirmée par le fait que ce sont les femelles de cette espèce qui quittent en plus grand nombre les enceintes expérimentales où l'intensité lumineuse interne est la plus faible (32 femelles E. orientalis piégées à la sortie des enceintes, pour seulement 3 femelles E. vuilleti). En quittant un habitat peu favorable, les femelles E. orientalis retrouvent toutes les potentialités d'une espèce généraliste apte à parasiter une gamme de 21 hôtes inféodés à des habitats aussi différents que ceux offerts par les forêts, savanes et cultures de l'Afrique de l'Ouest (Rasplus, 1988).
24Cependant les femelles, piégées à la sortie des enceintes ou trouvées à l'intérieur à la fin des 15 jours d'observation, ont parasité une partie des hôtes sains présents dans les enceintes. La présence de descendants femelles E. vuilleti et E. orientalis signifie que les femelles qui ont parasité les hôtes ont bien été accouplées par les mâles nés dans les enceintes (Damiens et al., 2002). Les effectifs des adultes nés à partir des hôtes parasités dans les enceintes sont significativement plus élevés dans les enceintes où l'intensité lumineuse est la plus faible. Les femelles E. vuilleti étant significativement engendrées en plus grand nombre, cela confirme le fait de leur plus grande activité dans l'obscurité (Ndoutoume et al., 2006).
25Le modèle expérimental prenant en compte l'intensité de la lumière dans les systèmes de stockage a permis de confirmer l'hypothèse selon laquelle les femelles de l'espèce généraliste E. orientalis s'évadent des structures de stockage. De ce fait, elles libèrent les bruches de leur pression parasitaire, ce qui favorise le maintien de ces ravageurs dans les greniers à des taux indésirables pour les agriculteurs. L'amélioration de la qualité des systèmes de stockage, notamment par l'utilisation de matériaux transparents, pourrait permettre le maintien des parasitoïdes dans les stocks. La présence des femelles E. orientalis dans les greniers pendant toute la période de stockage serait une pression parasitaire importante permettant de diminuer la densité des bruches.
Bibliographie
Alzouma I., 1987. Reproduction et développement de Bruchidus atrolineatus (Pic) aux dépens des cultures de Vigna unguiculata (Walp.) dans un agrosystème sahélien au Niger. Thèse de doctorat : Université de Tours (France).
Caswell G.H., 1974. The development and extension of non chemical control techniques for stored cowpeas in Nigeria. In: Brady, E.U. et al., eds. Proceedings of the first international working conference on stored-product entomology, October 7-11 1974, Savannah, Georgia, USA. Savannah, Georgia, USA: IWCSPP (International Working Conferences on Stored-Product Protection), 63-67.
Cortesero A.M., Monge J.P. & Monge J., 1997. Dispersal and parasitizing abilities of Eupelmus vuilleti (Hymenoptera: Eupelmidae), within a column of cowpea seeds. Env. Entomol., 26, 1025-1030.
Damiens D., Bressac C., Brillard J.P. & Chevrier C., 2002. Qualitative aspects of sperm stock in males and females from Eupelmus orientalis and Dinarmus basalis (Hymenoptera: Chalcidoidea), as revealed by dual fluorescence. Physiol. Entomol., 27, 97-102.
Doury G. & Rojas-Rousse D., 1994. Reproductive potential in the parasitoid Eupelmus orientalis (Hymenoptera: Eupelmidae). Bull. Entomol. Res., 84, 199-206.
Fleury F., Allemand R., Fouillet P. & Bouletreau M., 1995. Genetic variation in locomotor activity rhythm among populations of Leptopilina heterotoma (Hymenoptera: Eucoilidae) a larval parasitoid of Drosophila species. Behav. Genet., 25, 81-89.
Germain J.F., 1988. La diapause imaginale chez Bruchidus atrolineatus (Pic) en zone soudano-sahélienne. Thèse de doctorat : Université de Tours (France).
Huignard J., Leroi B., Alzouma I. & Germain J.F., 1985. Oviposition and development of Bruchidius atrolineatus (Pic) and Callosobruchus maculatus (Fab.) (Coleoptera: Bruchidae) in Vigna unguiculata Walp. cultures in Niger. Insect Sci. Appl., 6, 691-699.
Idi A., 1994. Suivi de l'évolution de la population des bruches (Bruchidus atrolineatus F., Coleoptera : Bruchidae) et de leurs parasitoïdes (Dinarmus basalis Rond, Hymenoptera : Pteromalidae ; Eupelmus vuilleti CRW, Hymenoptera : Eupelmidae ; Uscana lariophaga, Hymenoptera : Tricogrammatidae) dans les systèmes de stockage traditionnel de niébé (Vigna unguiculata Walp.) au Niger. Thèse de spécialité : Université de Niamey (Niger).
Jackai L.E.N. & Daoust R.A., 1986. Insect pests of cowpea. Annu. Rev. Entomol., 31, 95-119.
Kalmès R., Dronnet S., Rojas-Rousse D. & Chevrier C., 2002. Bilan nutritionnel et sex-ratio à l'émergence chez deux ectoparasitoïdes solitaires (Eupelmus vuilleti et Eupelmus orientalis Craw) (Hyménoptères : Eupelmidae) : comparaison lors du passage du parasitisme primaire (hôte primaire : Callosobruchus maculatus : Coléoptère, Bruchidae) à l'hyperparsitisme (hôte secondaire : Dinarmus basalis, Hyménoptère, Pteromalidae). In : Vème Conférence Internationale Francophone d'Entomologie, Québec, 14-18 juillet. Montréal : Université du Québec, 135.
Lévêque L. et al., 1993. Analysis of multiparasitism by Eupelmus vuilleti (Craw) (Eupelmidae) and Dinarmus basalis (Rondani) (Pteromalidae) in the presence of one of their commun hosts, Bruchidius atrolineatus (Pic) (Coleoptera: Bruchidae). Oecologia, 94, 272-273.
Monge J.P. & Huignard J., 1991. Population fluctuations of two bruchid species Callosobruchus maculatus (Fabricius) and Bruchidus atrolineatus (Pic) (Coleoptera: Bruchidae) and their parasitoids Dinarmus basalis (Randoni) and Eupelmus vuilleti (Crawford) (Hymenoptera: Eupelmidae) in a storage situation in Niger. J. Afr. Zool., 105, 187-196.
Monge J.P., Dupont P., Idi A. & Huignard J., 1995. The consequences of interspecific competition between Dinarmus basalis (Rond) (Hymenoptera: Pteromalidae) and Eupelmus vuilleti (CRW) (Hymenoptera: Eupelmidae) on the development of their host populations. Acta Oecol., 16(1), 19-30.
Ndoutoume A., 1996. Capacités parasitaires et plasticité comportementale de deux hyménoptères Eupelmidae (Eupelmus orientalis et Eupelmus vuilleti) partenaires de la communauté parasitaire des stades larvaires et nymphaux de Callosobruchus maculatus (Coléoptère Bruchidae). Thèse de doctorat : Université de Tours (France).
Ndoutoume A., Kalmès R. & Rojas-Rousse D., 2000. Reproductive potential of Eupelmus orientalis (Crawford) and Eupelmus vuilleti (Crawford) (Hymenoptera: Eupelmidae), two parasitoids of Bruchidae (Coleoptera) during the harvest and storage of cowpea pods (Vigna unguiculata L. Walp.). Afr. Entomol., 8, 201-209.
Ndoutoume A., Rojas-Rousse D. & Allemand R., 2006. Rythmes d'activité locomotrice de deux insectes parasitoïdes sympatriques : Eupelmus orientalis et Eupelmus vuilleti (Hyménoptère, Eupelmidae), C. R. Biol., 329, 476-482.
Rasplus J.-Y., 1988. La communauté parasitaire des Coléoptères séminivores des légumineuses dans une mosaïque forêt-savane en Afrique de l'Ouest (Lamto, Côte d'Ivoire). Thèse de doctorat : Université d'Orsay-Paris XI (France).
Rojas-Rousse D., Ndoutoume A. & Kalmès R., 1999. Hyperparasitisme facultatif de parasitoïdes en cours de développement par les femelles des ectoparasitoïdes Eupelmus vuilleti et Eupelmus orientalis Craw. C. R. Biol., 322, 393-399.
Rojas-Rousse D. et al., 2005. Capacité de reproduction des femelles Eupelmus orientalis (Eupelmidae), inséminées par des mâles developpés en hyperparasitoïdes de Dinarmus basalis (Pteromalidae). C. R. Biol., 328, 802-811.
Terrasse C. & Rojas-Rousse D., 1986. Distribution de la ponte et évitement du superparasitisme chez l'hyménoptère solitaire Bruchocida vuilleti (Hym., Eupelmidae), parasite des stades larvaires de son hôte, Callosobruchus maculatus F. (Coléoptère : Bruchidae). Z. Angew. Entomol., 101, 243-253.
Pour citer cet article
A propos de : Auguste Ndoutoume-Ndong
Ecole normale supérieure de Libreville. B.P. 17009. G-Libreville (Gabon). E-mail : augustendoutoume@caramail.com
A propos de : Danielle Rojas-Rousse
Université de Tours. Faculté des Sciences. Institut de Recherche sur la Biologie de l'Insecte. UMR-CNRS 6035. Parc de Grandmont. F-37200 Tours (France).