


- Accueil
- volume 16 (2012)
- numéro 3
- Impact of sowing density and nitrogen fertilization on Rumex obtusifolius L. development in organic winter cereal crops


Visualisation(s): 1473 (24 ULiège)
Téléchargement(s): 77 (2 ULiège)

Impact of sowing density and nitrogen fertilization on Rumex obtusifolius L. development in organic winter cereal crops

Notes de la rédaction
Received on December 17, 2010; accepted on January 12, 2012
Résumé
Impact de la densité de semis et de la fertilisation azotée sur le développement de Rumex obtusifolius L. en cultures de céréales d’hiver conduites en agriculture biologique. La gestion de la problématique posée par le Rumex obtusifolius L. (rumex à feuilles obtuses) est très importante en agriculture biologique. En effet, il y représente l’un des principaux facteurs limitant l’adhésion des éleveurs conventionnels à ce mode de production. C’est dans ce contexte que l’impact de deux facteurs, à savoir la densité de semis (SD) et le niveau de fertilisation azoté au tallage, sur le développement des populations de R. obtusifolius en cultures de céréales d’hiver, a été étudié. Les deux céréales d’hiver qui ont été considérées sont l’épeautre (Triticum spelta [L.] Thell.) et le triticale (×Triticosecale [A.Camus] Wittm.). Les résultats obtenus soulignent qu’une augmentation de la densité de semis et de la fertilisation azotée améliore la croissance de la culture et assure, ainsi, une meilleure couverture du sol à la fin du printemps, comme démontré par la diminution significative des radiations photosynthétiquement actives (PAR) qui arrivent au sol. Néanmoins, ces effets sont trop faibles, en avril, que pour limiter le développement des populations de R. obtusifolius suite au développement de nouvelles plantules. Une augmentation de la densité de R. obtusifolius parallèlement à l’augmentation du niveau de fertilisation azotée peut également être soulignée, ce qui illustre le caractère nitrophile de cette adventice. Bien qu’une augmentation de la densité de semis et du niveau de fertilisation azoté au tallage aient permis d’accroître significativement la densité du couvert végétal, ces deux techniques n’ont pas permis de réduire la densité de R. obtusifolius dans les cultures étudiées. Cependant, les rendements des céréales furent maintenus, voire améliorés. Cela souligne que, même en combinant les méthodes de luttes culturales testées et un désherbage mécanique, cette adventice pérenne est difficile à contrôler.
Abstract
The control of Rumex obtusifolius L. (broad-leafed dock) is very important in organic farming systems. Indeed, concerns about managing this weed without the use of herbicides is one of the major factors limiting the uptake of these systems by conventional farmers. Against this background, we analyzed the impact of two management practices on the development of R. obtusifolius populations in two winter cereal trials: spelt (Triticum spelta [L.] Thell.) and triticale (×Triticosecale [A.Camus] Wittm.). The management factors were sowing density (SD) and nitrogen fertilization (NF) at the tillering stage. The results showed that an increase in SD and NF led to stronger crop growth and better soil coverage by the end of spring, demonstrated by a significant decrease in photosynthetic active radiation (PAR) at soil level. However, although there was an SD effect, it was too weak in April to restrict an increase in R. obtusifolius populations through the recruitment of new R. obtusifolius plants. An increase in R. obtusifolius population density was also linked to an increase in the NF level, illustrating the nitrophilic character of this weed. Although an increase in SD and NF at the tillering stage led to a higher canopy density, these two practices failed to reduce R. obtusifolius density in the cereal crops. Nevertheless, cereal yields were shown to be maintained or improved. Our results indicate that, even when combining weed harrowing and some cultural weed control methods, this perennial weed is difficult to control.
Table des matières
1. Introduction
1The control of Rumex obtusifolius L. (broad-leafed dock), one of the five most widely distributed non-cultivated plant species in the world (Allard, 1965), is very important in organic farming systems, where concern about the difficulty of managing the plant without using herbicides is one of the major factors limiting the uptake of these systems by conventional farmers (Turner et al., 2007). Rumex obtusifolius is highly competitive, as it can flower several times a year, producing several thousand seeds. Moreover, it can regenerate from small fragments of the upper parts of its roots.
2As in the case of many perennial weeds, the long-term objective of control measures is not to eradicate this weed (Turner et al., 2007), but to limit its vigor and regrowth capacity where it has become well established (Stilmant et al., 2010), to hinder its seed production capacity and to reduce the emergence and establishment of new individuals (Zaller, 2004). To achieve this in an organic farming context, an integrated approach based on crop rotation is needed (Turner et al., 2007). The approach also needs to take into account several factors aiming to improve crop competitiveness and the ability of the crops to suppress weeds (O’Donovan et al., 2007). Among these factors, crop fertilization management (Zaller, 2004; Blackshaw et al., 2005; O’Donovan et al., 2007) or the adjustment of crop sowing density (O’Donovan et al., 2007; Zhao et al., 2007; Chauhan et al., 2011) are of special interest.
3In line with this approach, the aim of our research was to analyze the impact of sowing density (SD) and nitrogen fertilization (NF) at the tillering stage on R. obtusifolius population development in winter cereals. An increase in both SD and NF was tested in order to increase the rate and speed of soil cover by the cereal crops and thus to increase their competitiveness (O’Donovan et al., 2007) and their weed suppression ability (Murphy et al., 2008).
2. Materials and methods
2.1. Experimental site and design
4The impact of SD and NF at the tillering stage was tested during the 2005-2006 cropping season for two winter cereal species: spelt (Triticum spelta [L.] Thell.) and triticale (×Triticosecale [A.Camus] Wittm.).
5The trials were set up at Libramont (49°55’N – 5°35’E; 450 asl; 1,100 mm mean annual precipitation; 7.2 °C mean annual temperature). Daily average temperature, rainfall and global radiation were recorded in a meteorological station located at 500 m from these trials (www.pameseb.be). The trials were set up on a superficial loamy-stony soil in a field that had been farmed organically since 1998 and that was characterized by important R. obtusifolius occurrence. There were two independent trials, one for each cereal species, with four replications, set up in a fully randomized block design. Individual plots covered 15 m² (1.5 x 10 m).
2.2. Modalities and crop management sequences
6The three SDs tested for each cereal species were: standard density; standard density + 20% and standard density + 40%. This led to densities of 180, 220 and 260 kg·ha-1 for triticale and of 200, 240 and 280 kg·ha-1 for spelt.
7The three NF levels, applied at the end of the tillering stage were 0, 40 and 80 kg N·ha-1.
8The previous crops in the experimental field had been potatoes and spelt, respectively, for the spelt and triticale trials. An initial fertilization of 10 t of composted manure (6 kg N, 0.83 kg P and 3.73 kg K·t-1 of fresh material) was spread on the experimental field before plowing. The trials were sown on 18 October 2005 with inter-rows of 15 and 20 cm for spelt and triticale, respectively. The cereal varieties were COSMOS and BIENVENU for spelt and triticale, respectively. The organic seeds used had had no prior treatment. Weeding was carried out via double harrowing with a 3 m-wide light harrow (Einböck GmbH), on 11 April 2006. Differentiated fertilization was applied on 27 April 2006, in the form of chicken droppings. This organic fertilizer gave a dose of 40 kg N·t-1, 13 kg P·t-1 and 17 kg K·t-1.
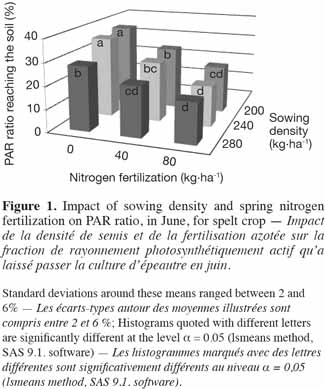
2.3. Recorded parameters
9Within each plot, the recorded parameters were:
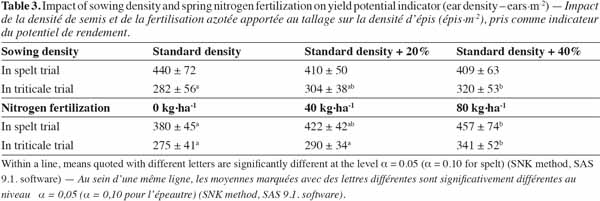
101. number of R. obtusifolius plants;
112. ratio of photosynthetically active radiation passing through the cereal crop canopy (Dingkuhn et al., 1999);
123. crop height (Dingkuhn et al., 1999);
134. ear density, determined by counting the ears in four rows of 0.5 m in each plot on 29 June.
14Parameters (2) and (3) gave an indication of crop competitiveness and hence weed suppression ability (Murphy et al., 2008) under the different management schemes. Parameter (4) indicated the potential yield. Grain yield was also recorded for triticale, but not for spelt because the crop was destroyed by boars before harvest.
15The density of R. obtusifolius was determined on three dates (Table 1). Seedlings were differentiated from plants regenerating from old rooting systems, based on cotyledon occurrence. The analyses focused on the total number of R. obtusifolius plants. The first count was performed on 4 April, before the NF application, in order to evaluate the impact of SD on this parameter.

16The PAR ratio is the ratio between the radiation, active from a photosynthetic perspective (400-700 nm), recorded below the crop canopy at soil level and above the crop canopy. These radiation measurements were recorded with a ceptometer (SunScan Canopy Analysis System). These measurements need to be carried out when there is a clear blue sky. So, because of the poor weather conditions in May 2006, the PAR ratio was recorded only twice: on 28 April, before there was likely to be any impact from the different fertilization schemes, as chicken droppings were spread on 27 April 2006, and on 6 June. On these dates, two measurements were performed in each plot, one at each extremity. In addition to the PAR ratio, crop height was recorded twice (Table 1).
2.4. Data analysis
17The density of R. obtusifolius observed in April was submitted to a 2-way mixed ANOVA analysis, which included, as factors, SD (three levels – fixed) and blocks (four levels – random). As there was a significant correlation between the densities in the subsequent counts and the initial R. obtusifolius density in April, the densities observed in May and June were submitted to a 3-way mixed CO-VARIANCE analysis, with the April density used as a co-variable. The other parameters were tested using the 3-way mixed ANOVA model, which included, as factors, SD (three levels – fixed), NF in spring (three levels – fixed) and blocks (four levels – random). The fixed factors and their interaction were tested against their interactions with the blocks.
18All the analyses were performed using the GLM procedure from the SAS software (SAS 9.1. software – SAS Inc, Cary, NC, USA). Comparisons of multiple means were performed using the Student-Newman-Keuls (SNK) method.
3. Results
19With a measurement of 556 mm rainfall by the end of May, 2006 had a similar cumulated rainfall level to the average value recorded over 2001-2011 (552 mm). Nevertheless, there was a deficit of 52% during the winter period with 154 mm instead of 321 mm on average, while spring was characterized by a rainfall excess of 47% (427 mm instead of 290 mm). The average temperature in 2006 was, by the end of May, 4.0 °C as against the 5.4 °C recorded, for the same months, during the period 2001 to 2011. The observed deficit occurred during the winter period with an average temperature of -0.4 °C as against the 1.8 °C recorded during 2001-2011. Spring 2006, with an average temperature of 10.2 °C, was near the average of 10.6 °C. Excesses of global radiation, of 17 and 9%, respectively, during the winter and spring periods, were recorded in 2006. So 2006 was characterized by a cold, dry and sunny winter and a wet but sunny spring. These last conditions favored the germination of R. obtusifolius seeds.
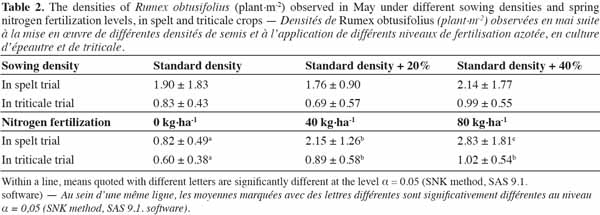
20Under these conditions, by the end of May, recorded R. obtusifolius densities were, on average, 1.94 ± 1.52 and 0.84 ± 0.52 plants per m2, respectively, in the spelt and the triticale trials.
3.1. R. obtusifolius densities at the end of winter
21In both trials, the R. obtusifolius densities recorded on 4 April were, on average, 0.11 ± 0.14 plants per m2, all were plants that had regenerated from old rooting systems. No significant impact of SD on this parameter was recorded in the spelt trial (F(2,24) = 1.18; p = 0.369), but there was a significant impact (F(2,24) = 8.48; p = 0.018) in the triticale trial. Here, the 260 kg·ha-1 sowing rate led to the highest R. obtusifolius density, with 0.18 plants per m2 as opposed to 0.08 plants per m2, on average, for the other two sowing rates.
22As, at this time, no gradient between R. obtusifolius density and SD was highlighted, for either the spelt or the triticale trials, and as no R. obtusifolius seedling was identified, we suggest that the effect recorded in the triticale trial was linked more closely to density heterogeneity in the initial R. obtusifolius population distribution than to the cereal SD. In order to manage this potential bias, covariance analyses using the densities recorded on 4 April as the co-variable were performed on subsequent counts. The correlation between the densities recorded in these subsequent counts and the density recorded on 4 April was always higher than 0.51 (N = 36; p < 0.01).
3.2. R. obtusifolius densities at the end of spring
23As the correlation between the densities recorded in May and June were very highly significant (rspelt = 0.89; N = 36; p < 0.001 - rtriticale = 0.55; N = 36; p < 0.001), we only analyzed the densities recorded in May because, during this month, counting was easier and therefore probably more accurate, the crop growth being lighter than in June.
24Whatever the cereal species, R. obtusifolius population density was positively and very highly significantly influenced by the NF (Fspelt(2,6) = 32.9; p < 0.001 - Ftriticale(2,6) = 24.6; p = 0.001), but not by the SD (Fspelt(2,6) = 0.4; p = 0.709 - Ftriticale(2,6) = 0.3; p = 0.758), without any interaction between these two factors (Fspelt(4,11) = 0.4; p = 0.799 - Ftriticale(4,11) = 0.9; p = 0.483). However, in both the spelt and triticale trials, the highest R. obtusifolius densities were observed under the highest SD, and the lowest weed densities were recorded under the intermediate SD (Table 2).
3.3. Crop canopy evolution
25The PAR ratio measured on 28 April 2006 revealed a marginally significant (FPAR ratio-spelt(2,6) = 4.6; p = 0.061) and a significant (FPAR ratio-triticale(2,6) = 6.1; p = 0.036) impact of SD on this parameter in the spelt and triticale crops, respectively. In spelt, the SD of 280 kg·ha-1 led to the highest PAR interception (36 ± 6%) compared with the SD of 200 and 240 kg·ha-1 (30 ± 4% and 29 ± 3%, respectively). In triticale, the SD of 220 and 260 kg·ha-1 led to significantly higher PAR interceptions (39 ± 4% and 38 ± 5%, respectively) than the SD of 180 kg·ha-1 (34 ± 5%).
26In spelt, in June, the PAR ratio reaching the soil was significantly influenced by the NF (FPAR ratio(2,6) = 78.1; p < 0.001) and the SD (FPAR ratio(2,6) = 5.2; p = 0.048) levels without interaction between these two factors (p > 0.10). There was an increase in PAR interception by the crop with an increase in both NF and SD (Figure 1). So the PAR ratios reaching the soil were 29 ± 7, 27 ± 9 and 23 ± 6%, respectively, for SDs of 200, 240 and 280 kg·ha-1. These ratios were 34 ± 5, 26 ± 5 and 19 ± 4%, respectively, for NF of 0, 40 and 80 kg·ha-1. Crop height, with an average value of 84.3 ± 5.0 cm, was significantly impacted by NF (Fcrop height(2,6) = 10.6; p = 0.011) but not by SD (Fcrop height(2,6) = 2.0; p = 0.214) levels. The canopy height obtained following the application of 40 or 80 kg N·ha-1 was, at 85.9 ± 3.7 cm, higher than the one observed without nitrogen fertilization (81.2 ± 5.9 cm).
27In triticale, canopy height (109.8 ± 3.4 cm) was not impacted either by NF (Fcrop height(2,6) = 0.6; p = 0.567) or by SD (Fcrop height(2,6) = 1.9; p = 0.229). The PAR ratio reaching the soil was significantly and negatively impacted by SD (FPAR ratio(2,6) = 13.3; p = 0.006). The ratio was 26 ± 6, 24 ± 4 and 22 ± 4% for SDs of, respectively, 180, 220 and 260 kg·ha-1. This ratio was not significantly modified by the level of NF (FPAR ratio(2,6) = 3.1; p = 0.118) even though the ratio showed a trend of decreasing as NF increased. The ratio was 27 ± 6, 24 ± 4 and 21 ± 4% for NF of, respectively, 0, 40 and 80 kg·ha-1. As for spelt, there was no significant interaction between the SD and NF factors (p > 0.10).
3.4. Yield indicators
28A significant interaction between SD and NF was observed for ear density in the spelt crop (F(4,12) = 6.5; p = 0.005). The impact of NF (F(2,6) = 4.6; p = 0.062) was modulated by SD: an application of 80 kg·ha-1 led to an increase in ear density for the SD of 200 and 280 kg·ha-1, while an application of 40 kg·ha-1 led to an increase in ear density at the SD of 240 and 280 kg·ha-1.
29Triticale ear density was significantly and positively affected by both SD (F(2,6) = 7.3; p = 0.025) and NF (F(2,6) = 6.7; p = 0.030), without interaction between these two factors. The SD of 260 kg·ha-1 led to a 13% increase in ear density compared with the SD of 180 kg·ha-1. The application of 80 kg N·ha-1 led to an increase in triticale ear density of 24% compared to the plots that received no nitrogen (Table 3). With an average of 4.0 ± 0.5 t·ha-1, triticale grain yield was not significantly affected by SD (F(2,6) = 2.0; p = 0.220), but it was affected by NF (F(2,6) = 28.3; p = 0.001): the application of 80 kg N·ha-1 in spring, in the form of chicken droppings, led, with 4.5 ± 0.2 t·ha-1, to a yield increase of 0.9 t·ha-1, which was 25% higher than in the plots that received no nitrogen (3.6 ± 0.4 t·ha-1). The application of an intermediate NF of 40 kg·ha-1 led to an intermediate and significantly different yield of 4.0 ± 0.3 t·ha-1. As for triticale ear density, the interactions between SD and NF were not significant (p > 0.10) in explaining triticale grain yield variations.
4. Discussion and conclusion
30As indicated by the decrease in the PAR ratio, at soil level, an increase in both SD and NF led to stronger crop development and better soil cover by the end of spring. This effect, even where there was an SD impact in April, was too weak to restrict the development of R. obtusifolius populations through the recruitment of new individual plants from the germination of seeds able to survive and accumulate for several years in the soil (Zaller, 2004). In each trial, this was shown by the significant correlation observed between the densities recorded later in spring and the density of R. obtusifolius regenerated from old rooting systems and recorded on 4 April. Nevertheless, the multiplication factors of 14 and 3 in the spelt and triticale trials, respectively, observed between the April and June counts, were very different. This can be explained by less efficient control of R. obtusifolius in the organic potato crop, preceding the spelt trial, than in the organic spelt crop, preceding the triticale trial (Hatcher et al., 2003). These results stress the need to control weed infestation throughout a crop rotation in order to reduce seed reserves in the soil. This is especially true in organic farming (Turner et al., 2007), where the repeated chemical treatments, often needed in conventional systems to control R. obtusifolius (Niggli et al., 1993), are not allowed.
31In their study, Zhao et al. (2007) found they needed to raise the aerobic seeding rate from 100 to 300 viable seeds per m2 in order to observe a significant impact of the crop on weed suppression ability. In comparison, the increase of our standard seedling rate by 40% could have been too weak.
32Achieving good soil cover by a cereal crop in spring in order to reduce the recruitment of new R. obtusifolius plants is very important because during the season, R. obtusifolius seeds gradually lose dormancy (Benvenuti et al., 2001). Their germination is stimulated by repeated rapid heating (Van Assche et al., 1997) or, at lower temperatures, by more light (Benvenuti et al., 2001), two conditions that in our trials occurred more often on bare soil than on shaded soil. In a trial focusing on the development of the R. obtusifolius population during grassland implantation, it was found that, under good conditions, about 85% of this weed’s seeds emerged within 3 months of sowing (Hongo, 1989).
33In the present study, the lack of significant impact of NF on the PAR ratio recorded under the triticale crop raises questions, as this factor led to a significant increase in the triticale yield. One explanation could be found in the level of crop development when this parameter was measured, as underlined by the height difference between the crops, favoring the triticale crop (109.8 cm as against 84.3 cm for spelt). Such a level of development would have limited PAR variation at soil level. An earlier measurement in triticale would have increased the sensitivity of this parameter. However, the good conditions necessary to perform these measures (clear blue sky) were not available at the time.
34In order to achieve good cover, apart from increasing SD (for a review, see O’Donovan et al., 2007) and stimulating cover development through N application, cereal breeders aim to select varieties with good weed suppression ability traits (Wicks et al., 2004; O’Donovan et al., 2007). Among the desirable traits are large seedling ground cover, plant height, canopy structure and a large leaf area (Murphy et al., 2008). With the exception of plant height, no information has previously been available to enable farmers to make a choice between the spelt and triticale varieties available for organic farming. With the intention of being able to use the results of these trials to help farmers make an informed choice between spelt and triticale varieties, we used the most widely used varieties found in organic fields.
35While the tested strategy can restrict weed seedling recruitment by limiting seed germination, it seems to have no impact on the regrowth of well established R. obtusifolius, although it can modulate its reproductive capacity (Niggli et al., 1993). Nor does the strategy have an impact on R. obtusifolius plantlets, which increase their petiole and leaf area development in response to shaded surroundings due to crop competition (Jeangros et al., 1990). Nevertheless, this morphological flexibility has its limits. Thus, Hongo (1989) showed that R. obtusifolius survival is enhanced by frequent cutting, at least during its second growing season.
36Jeangros et al. (1990) suggest that competition for N is the main factor limiting the initial development of R. obtusifolius. This is supported by our results. We observed an increase in R. obtusifolius population density linked to the level of organic NF, although this factor is correlated to a stronger crop growth and higher yields. This illustrates the nitrophilic characteristic of this weed (Niggli et al., 1993; Zaller, 2004; Stilmant et al., 2007).
37The lack of control of R. obtusifolius seedling development in these trials could also be linked to their emergence after weed harrowing, performed on 11 April, in order to control other weed species at an early growth stage (Hatcher et al., 2003). This practice could have stimulated R. obtusifolius seed germination by moving the seeds towards the soil surface (Benvenuti et al., 2001). As a result, in order to control weed growth, it would have been useful to carry out an additional weed harrowing two weeks later, at the end of April (Hatcher et al., 2003). This was not done because of the poor weather conditions.
38In conclusion, although an increase in SD and NF at the tillering stage led to a higher canopy density, these two practices failed to significantly reduce R. obtusifolius density in these winter cereal crops. Nevertheless, cereal yields were maintained or improved. Our results regarding the lack of suppression of R. obtusifolius support earlier findings that, even when combining mechanical and cultural weed control methods, this weed is difficult to control (Hatcher et al., 2003; Zaller, 2004). As stressed by Zaller (2004) and illustrated by the highest R. obtusifolius densities occurring in spelt plots after a potato crop, it is necessary to implement mechanical and cultural weed control strategies (e.g. crop rotation design, soil cultivation methods, organic fertilizer composting) throughout the crop rotation in order to control this perennial weed. Further work, however, is needed to design such long-term control strategies (Zaller, 2004).
39Abbreviations
40SD: sowing density
41NF: nitrogen fertilization
42PAR: photosynthetic active radiation
43Acknowledgements
44The authors thank Mr J. Herman for his technical support and the Walloon Agricultural Research Centre for funding.
Bibliographie
Allard R.W., 1965. Genetic systems associated with colonizing ability in predominantly self-pollinating species. In: Bakr H.G. & Stebbins G.L., eds. The genetics of colonizing species. New York, USA: Academic Press, 49-75.
Benvenuti S., Macchia M. & Miele S., 2001. Light, temperature and burial depth effects on Rumex obtusifolius seed germination and emergence. Weed Res., 41, 177-186.
Blackshaw R.E., Molnar L.J. & Larney F.J., 2005. Fertilizer, manure and compost effects on weed growth and competition with winter wheat in western Canada. Crop Prot., 24, 971-980.
Chauhan B.S., Singh V.P., Kumar A. & Johnson D.E., 2011. Relations of rice seeding rates to crop and weed growth in aerobic rice. Field Crops Res., 121, 105-115.
Dingkuhn M., Johnson D.E., Sow A. & Audebert A.Y., 1999. Relationships between upland rice canopy characteristics and weed competitiveness. Field Crops Res., 61, 79-95.
Hatcher P.E. & Melander B., 2003. Combining physical, cultural and biological methods: prospects for integrated non-chemical weed management strategies. Weed Res., 43, 303-322.
Hongo A., 1989. Survival and growth of seedlings of Rumex obtusifolius L. and Rumex crispus L. in newly sown grassland. Weed Res., 29, 7-12.
Jeangros B. & Nösberger J., 1990. Effects of an established sward of Lolium perenne L. on the growth and development of Rumex obtusifolius L. seedlings. Grass Forage Sci., 45, 1-7.
Murphy K.M., Dawson J.C. & Jones S.S., 2008. Relationship among phenotypic growth traits, yield and weed suppression in spring wheat landraces and modern cultivars. Field Crop Res., 105, 107-115.
Niggli U., Nösberger J. & Lehmann J., 1993. Effects of nitrogen fertilization and cutting frequency on the competitive ability and the regrowth capacity of Rumex obtusifolius L. in several grass swards. Weed Res., 33, 131-137.
O’Donovan J.T. et al., 2007. Integrated approaches to managing weeds in spring-sown crops in western Canada. Crop Prot., 26, 390-398.
Stilmant D. et al., 2007. Le rumex à feuilles obtuses dans les systèmes herbagers : importance de la problématique, lutte chimique et méthodes alternatives. Fourrages, 192, 477-493.
Stilmant D., Bodson B., Vrancken C. & Losseau C., 2010. Impact of cutting frequency on the vigour of Rumex obtusifolius. Grass Forage Sci., 65, 147-153.
Turner R.J. et al., 2007. Organic weed management: a review of the current UK farmer perspective. Crop Prot., 26, 377-382.
Van Assche J.A. & Van Nerum D.M., 1997. The influence of the rate of temperature change on the activation of dormant seeds of Rumex obtusifolius L. Funct. Ecol., 11, 729-734.
Wicks G.A. et al., 2004. Winter wheat cultivar characteristics affect annual weed suppression. Weed Technol., 18, 988-998.
Zaller J.G., 2004. Ecology and non-chemical control of Rumex crispus and R. obtusifolius (Polygonaceae): a review. Weed Res., 44, 414-432.
Zhao D.L., Bastiaans L., Altin G.N. & Spiertz J.H.J., 2007. Interaction of genotype x management on vegetative growth and weed suppression of aerobic rice. Field Crops Res., 100, 327-340.
Pour citer cet article
A propos de : Didier Stilmant
Walloon Agricultural Research Centre. Farming Systems, Territories and Information Technology Unit. Rue du Serpont, 100. B-6800 Libramont (Belgium). E-mail: stilmant@cra.wallonie.be
A propos de : Bernard Bodson
Univ. Liege - Gembloux Agro-Bio Tech. Unit of Plant Science of the moderate Regions. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
A propos de : Céline Losseau
Univ. Liege - Gembloux Agro-Bio Tech. Unit of Plant Science of the moderate Regions. Passage des Déportés, 2. B-5030 Gembloux (Belgium).