


- Accueil
- Volume 11 (2007)
- numéro 3
- Effet de la salinité sur la répartition des cations (Na+, K+ et Ca2+) et du chlore (Cl-) dans les parties aériennes et les racines du ray-grass anglais et du chiendent


Visualisation(s): 12862 (86 ULiège)
Téléchargement(s): 619 (9 ULiège)

Effet de la salinité sur la répartition des cations (Na+, K+ et Ca2+) et du chlore (Cl-) dans les parties aériennes et les racines du ray-grass anglais et du chiendent

Notes de la rédaction
Reçu le 4 janvier 2005, accepté le 5 décembre 2006.
Résumé
Des plantes de chiendent, variété Tifway, et de ray-grass anglais, variété Perfect, issues de semis ont été cultivées en plaques de végétation et arrosées à l’eau courante additionnée de différentes doses de NaCl (50 à 150 mM). Une culture a été menée pendant la période estivale et une autre pendant la période hivernale. Les caractéristiques ioniques des organes végétatifs des plantes (feuilles, racines) dépendent de l’espèce et de la saison de culture. Les plantes de ray-grass anglais et de chiendent ont accumulé du Na+ et Cl- dans leurs tissus mais les teneurs foliaires en ces ions ont été généralement plus élevées chez le ray-grass anglais surtout sur la forte concentration de NaCl. La salinité a entraîné des modifications des teneurs en K+ et Ca2+ selon l’espèce, la nature de l’organe et la saison.
Abstract
Salinity effect on the assessment of cations (Na+, K+ and Ca2+) and chlorine (Cl-) in the aerial parts and roots of perennial ryegrass and bermudagrass. Seedling plants of bermudagrass, var. Tifway, and perennial ryegrass, var. Perfect, were cultivated in growth plate and sprinkled with running water added with different contents of NaCl (50 to 150 mM). One cultivation is led during summer time and another one during winter time. The ionic characteristics of vegetative organs of plants (leaves, roots) depend of the species and the season of cultivation. The plants of perennial ryegrass and bermudagrass accumulated Na+ and Cl- in their tissues but leaf contents for these ions were generally higher in perennial ryegrass especially at the highest concentration of NaCl. Salinity led to modifications in K+ and Ca2+ contents according to the species, kind of organ and season.
Table des matières
1. Introduction
1Le chiendent et le ray-grass anglais sont deux graminées très utilisées comme espèces gazonnantes. Elles entrent, en effet, dans la composition de nombreux mélanges, notamment ceux des gazons de sport. Le comportement de ces deux espèces vis-à-vis de la salinité diffère. D’après Lemaire (1993), le ray-grass anglais est une espèce moyennement sensible à la salinité (tolérant une conductivité électrique CE de 4 à 8 mS.cm-1) alors que le chiendent est classé comme une espèce tolérante (supportant une CE de 8 à 16 mS.cm-1). D’autre part, la biologie des deux espèces diverge dans la mesure où le ray-grass anglais se confirme comme une espèce de saison froide alors que le chiendent est plutôt une espèce de saison chaude.
2Les effets nutritionnels de la salinité incluent les deux actions primaires du sel sur les plantes : la toxicité directe due à l’accumulation excessive des ions dans les tissus et un déséquilibre nutritionnel provoqué par l’excès de certains ions. Des concentrations salines trop fortes dans le milieu provoquent une altération de la nutrition minérale des plantes (Levigneron et al., 1995).
3L’accumulation des ions Na+ dans la plante limite l’absorption des cations indispensables tels que K+ et Ca2+. Il y aurait une compétition entre Na+ et Ca2+ pour les mêmes sites de fixation apoplasmique. L’interaction entre les ions Na+ et Ca2+ influe sur la croissance des racines d’orge (Heyder, Greenway, 1965 cités par Jendoubi, 1997).
4L’accumulation des ions Na+ affecte l’absorption de K+ et ceci en fonction de la concentration du premier élément, cependant, la présence de Na+ en faible concentration peut augmenter l’absorption de K+, tandis qu’une concentration élevée en Na+ diminue l’absorption de K+ chez le riz (Levitt, 1980) et la canne à sucre (Nimbalkar, Joshi, 1975). Cette absorption peut même s’arrêter complètement chez le haricot (Hamza, 1977) et le laurier rose (Hajji, 1980) cultivés en présence de chlorure de sodium (NaCl) à 12 g.l-1.
5Chez le blé, tout comme chez le riz et la canne à sucre, la concentration élevée de NaCl diminue également l’absorption de Ca2+. Chez Brassica campestris L., qui est relativement tolérante au sel, l’augmentation de la concentration en Na+ s’accompagne d’une réduction de la concentration en Mg, K, N, P et Ca dans la plante (Levitt, 1980). Ce déséquilibre nutritionnel est une cause possible des réductions de croissance en présence de sel lorsque des ions essentiels comme K+, Ca2+ ou NO3- deviennent limitants (Soltani, 1988).
6Selon Ehret et al. (1990), les céréales représentent les plantes les plus sensibles à l’insuffisance en Ca. Chez le triticale cultivé sous stress salin, le calcium est plus limitant que le potassium (Yakoubi-Tej, 1992).
7La présence de Cl- inhibe l’absorption de NO3- (Smith, 1973). Le chlorure diminue la concentration en phosphore dans les feuilles de blé (Piri, 1991).
8La protection contre la toxicité des ions se fait par plusieurs mécanismes qui diffèrent selon la catégorie de la plante. Chez les plantes sensibles à NaCl, Na s’accumule dans les racines mais il est exclu des feuilles (Slama, 1986). Ces plantes sont dites « excluder » vis-à-vis de Na. à l’inverse, les plantes tolérant NaCl sont dites « includer » car elles ont en général des feuilles plus chargées en Na que les racines lorsqu’elles sont cultivées en présence de sel (Slama, 1986).
9L’objectif du présent travail est d’étudier la nutrition minérale de deux graminées à gazon communément utilisées : le ray-grass anglais et le chiendent, en présence de différentes concentrations de NaCl. Par ailleurs, l’expérimentation a été menée en deux saisons de culture, l’hiver et l’été, pour mieux intégrer la biologie de l’espèce dans la caractérisation de leur tolérance à la salinité.
2. Matériel et méthodes
2.1. Matériel végétal
10Deux espèces de graminées sont utilisées : le ray-grass anglais (Lolium perenne L., variété Perfect) et le chiendent (Cynodon dactylon L., variété Tifway).
11Ces 2 espèces sont très employées en Tunisie pour l’engazonnement et le regarnissage des terrains de sport, en particulier les terrains de football et les golfs (départs et parcours).
2.2. Protocole expérimental
12Conduite des cultures. Les expérimentations sont réalisées à l’Institut Supérieur Agronomique de Chott Mariem, Sousse (Tunisie).
13La culture de ces 2 graminées est réalisée pendant 2 saisons de l’année : l’été et l’hiver, correspondant chacune à une période de croissance active ou ralentie pour l’une ou l’autre des 2 espèces.
14Culture estivale. Le semis des graines des 2 espèces est effectué le 15 mai 2003 en conteneurs en polystyrène de dimensions 60 x 40 x 7 cm, sur un substrat composé de terre de jardin, de fumier de ferme décomposé et de sable grossier à proportions égales.
15Les conteneurs de culture sont placés en plein air dans des conditions naturelles.
16Au début, l’arrosage des graines est effectué à l’eau courante et exécuté sous forme de fines gouttelettes pour éviter le déchaussement des graines. Cette eau renferme entre autres ions (en méq.l-1) : Na+ (10,0), K+ (0,2), Ca2+ (8,3) et Cl- (11,2). Une fois la levée réalisée et lorsque les pousses atteignent une hauteur de 3 cm environ, l’application des traitements salins commence et les arrosages s’effectuent ainsi à l’eau courante additionnée ou non de différentes doses de NaCl (50 à 150 mM). Le milieu témoin est arrosé uniquement à l’eau courante. Les arrosages sont effectués de manière à maintenir le substrat de culture à la capacité au champ. La quantité d’eau apportée lors de chaque arrosage est de 1,5 litre par conteneur.
17Le dispositif expérimental est réalisé en split-plot, en blocs aléatoires complets, avec 2 espèces (chiendent et ray-grass anglais) et 4 traitements salins (témoin, 50, 100 et 150 mM NaCl). Le nombre de blocs est de 3. Dans le bloc, chaque traitement est constitué d’un conteneur de végétation. Cette culture s’est poursuivie jusqu’au 15 octobre 2003.
18Culture hivernale. Le semis est réalisé le 26 novembre 2003 et la culture est maintenue jusqu’au 26 avril 2004. Dans ce cas, les conteneurs de végétation sont placées sous abri-serre plastique pour les protéger contre la pluie. Le protocole expérimental et la conduite des arrosages sont les mêmes que précédemment.
19Conditions de culture. Dans la région de Chott Mariem, Sousse (Tunisie), les températures moyennes enregistrées en plein air pour la culture estivale (mai-octobre 2003) ont oscillé entre 20,1 et 28,3 °C, celles enregistrées sous abri-serre plastique pour la culture hivernale (novembre 2003 - avril 2004) se sont situées entre 17,9 et 20,2 °C.
20Dosage des éléments minéraux. La composition chimique de la matière végétale est déterminée à l’arrêt de la culture soit après 4 mois de traitement salin. Les plantes sont séparées en parties aérienne et racinaire et leurs organes sont préalablement desséchés pendant 48 heures à 80 °C et mis dans des piluliers contenant 25 ml d’acide nitrique (0,1 N). Les différents dosages se font sur le filtrat de l’extraction acide à froid. Les cations Na+, K+, Ca2+ sont dosés par spectrophotométrie de flamme en émission (photomètre Jenway). L’ion Cl- est dosé par colorimétrie à l’aide d’un colorimètre (Sherwood). Les analyses minérales concernent 8 plantes prélevées au hasard dans chaque conteneur de culture, soit au total 192 plantes.
3. Résultats
3.1. Croissance pondérale des organes végétatifs
21Le rapport entre la biomasse sèche des racines et celle des feuilles est toujours inférieur à 1, ce qui montre que la masse de matière sèche des feuilles est plus élevée que celle des racines (Tableau 1). Ce paramètre augmente avec la salinité à l’exception du chiendent en culture estivale où ce rapport est remarquablement constant. Cela veut dire, qu’en présence de sel, la croissance pondérale des racines devient supérieure à celle des feuilles. En été et en dépit de la contrainte saline, le chiendent arrive à maintenir sa croissance foliaire au même rythme que la croissance des racines. De plus, en présence de fortes doses de NaCl (100 et 150 mM), ce rapport est plus faible en cette saison, ce qui signifie que, dans des conditions de chaleur et de salinité, la croissance des racines, chez cette espèce, est plus affectée que celle de la partie aérienne. Pour les 2 périodes de culture, ce rapport est nettement plus élevé chez le ray-grass anglais indiquant une partie racinaire plus développée chez cette espèce que la partie aérienne.
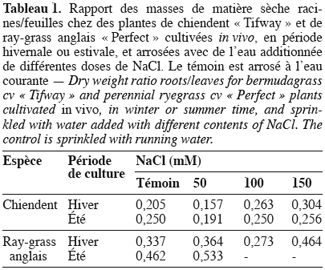
22Le sodium. Pendant la période hivernale, les teneurs en Na+ des feuilles et des racines de chiendent sont les plus faibles sur le milieu témoin et sont respectivement de 0,17 et 0,67 méq.g-1 de matière sèche (MS) (Figure 1). Ces teneurs augmentent de façon systématique avec la concentration de NaCl pour atteindre sur 150 mM les valeurs respectives de 0,70 et 1,11 méq.g-1 MS. Chez cette espèce, ce sont les racines qui se chargent remarquablement plus que les feuilles en Na+.
23Chez le ray-grass anglais, les teneurs les plus faibles sont obtenues sur le milieu témoin et sont comparables pour les racines et les feuilles (0,36 méq.g-1 MS). Il y a une élévation systématique des teneurs des organes en Na+ avec l’augmentation de la salinité de l’eau d’arrosage. Sur NaCl 150 mM, les charges foliaire et racinaire en Na+ s’élèvent respectivement à 1,56 et 1,12 méq.g-1 MS. Contrairement au chiendent, ce sont les feuilles qui accumulent plus de Na que les racines.
24Chez le chiendent cultivé en période estivale, les teneurs en Na+ des feuilles et des racines des plantes témoins sont comparables et faibles (0,13 méq.g-1 MS). Tous les organes de la plante accumulent cet ion en présence de NaCl, et les parties aériennes se chargent relativement plus que les racines en ce cation. Sur NaCl 150 mM, la teneur foliaire en Na+ atteint 0,61 méq.g-1 MS.
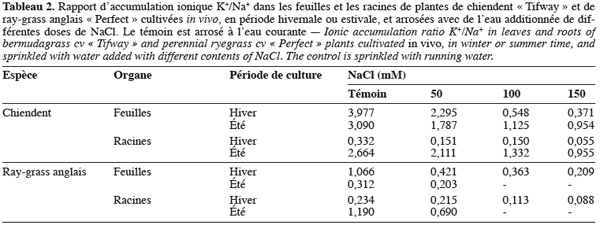
25Le ray-grass anglais a montré, en culture estivale, une grande sensibilité à la salinité et les plantes arrosées à l’eau additionnée de 100 et 150 mM NaCl ont totalement disparu. Pour cette raison, la composition minérale de ces plantes n’a pu se faire et seules les plantes témoins et cultivées en présence de NaCl 50 mM ont fait l’objet de cette analyse.
26Sur le milieu témoin, la charge tissulaire des feuilles et des racines en Na+ est comparable et se situe à 0,3 méq.g-1 MS. Sur NaCl 50 mM, ces organes s’enrichissent en Na+ et leurs teneurs atteignent respectivement 0,84 et 0,64 méq.g-1 MS.
27Le potassium. L’examen de la figure 2 montre que, chez le chiendent cultivé en saison hivernale, c’est la solution témoin d’arrosage qui autorise les teneurs en K+ les plus élevées et sont plus de 3 fois plus élevées dans les feuilles (0,70 méq.g-1 MS) que dans les racines (0,22 méq.g-1 MS). Le sel réduit nettement les teneurs dans ces organes lorsque sa concentration dépasse 50 mM NaCl. Sur NaCl 150 mM, les charges foliaire et racinaire en K+ tombent respectivement à 0,26 et 0,06 méq.g-1 MS.
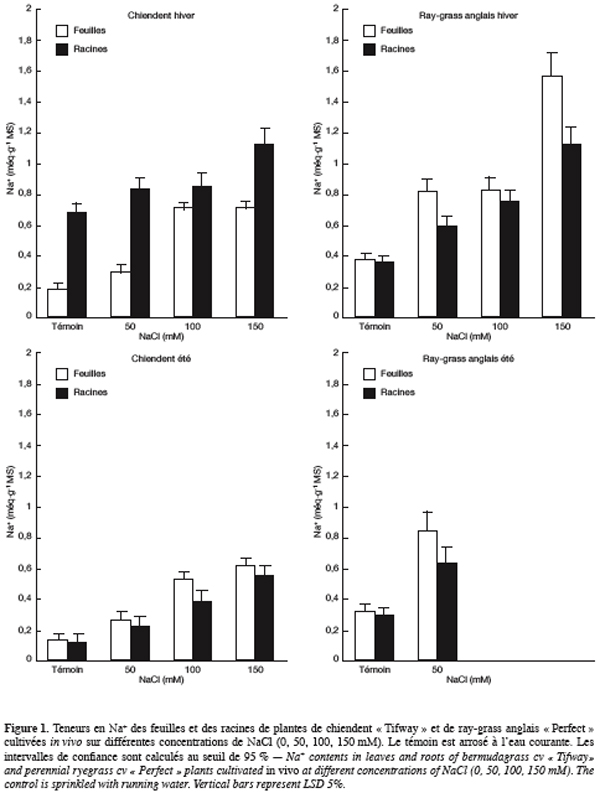
28Chez le ray-grass anglais, les teneurs foliaires les plus élevées en potassium sont obtenues sur le milieu témoin (0,40 méq.g-1 MS) et sont plus de 4 fois supérieures à celles des racines (0,085 méq.g-1 MS). La salinité de la solution d’arrosage entraîne une baisse discrète des teneurs foliaires en K+ alors que celles des racines paraissent insensibles au sel.
29En été, la teneur foliaire en K+ du chiendent témoin est de 0,41 méq.g-1 MS, celle des racines est de 0,33 méq.g-1 MS. En présence de NaCl, les teneurs en ce cation augmentent dans les feuilles et les racines pour atteindre respectivement 0,58 et 0,54 méq.g-1 MS sur NaCl 150 mM.
30Chez le ray-grass anglais, les teneurs des racines en K+ deviennent 2,5 à 3,5 fois supérieures à celles des feuilles, selon la dose de sel dans la solution d’arrosage. La salinité fait augmenter ces teneurs dans les feuilles et les racines qui passent respecti-vement de 0,10 et 0,35 méq.g-1 MS sur le milieu témoin à 0,17 et 0,44 méq.g-1 MS en présence de NaCl 50 mM.
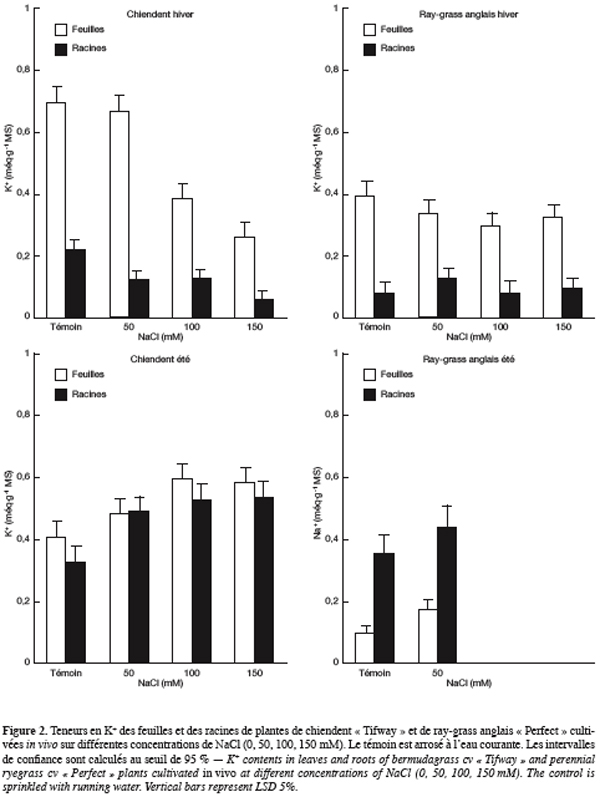
31Le calcium. En période hivernale, le chiendent cultivé sur le milieu témoin présente des teneurs foliaire et racinaire en Ca2+ de valeurs respectives 0,20 et 0,44 méq.g-1 MS (Figure 3). La charge des racines en calcium est supérieure à celle des feuilles mais les différences s’estompent à partir de NaCl 100 mM. Les teneurs des feuilles en Ca2+ sont peu modifiées par la salinité de l’eau d’arrosage alors que celles des racines diminuent nettement.
32Chez le ray-grass anglais arrosé avec la solution témoin, la teneur en calcium des racines (0,44 méq.g-1 MS) est supérieure à celle des feuilles (0,16 méq.g-1 MS). La situation s’inverse en présence de NaCl et les feuilles deviennent plus accumulatrices de calcium que les racines. La salinité charge de plus en plus les feuilles en Ca2+ alors qu’elle fait baisser les teneurs racinaires en cet ion. Sur NaCl 150 mM, les charges foliaire et racinaire en Ca2+ atteignent respectivement 0,56 et 0,09 méq.g-1 MS.
33En été, les teneurs des feuilles et des racines de chiendent en Ca2+ sont faibles et ne dépassent pas 0,2 méq.g-1 MS. La salinité provoque une augmentation discrète des teneurs foliaires en ce cation alors que cette augmentation est plus marquée dans le cas des racines. Sur NaCl 150 mM, les charges foliaire et racinaire en Ca2+ atteignent respectivement 0,08 et 0,17 méq.g-1 MS.
34Chez le ray-grass anglais arrosé avec la solution témoin, la teneur foliaire en Ca2+ est de 0,10 méq.g-1 MS, celle des racines est plus de 3 fois supérieure, soit 0,36 méq.g-1 MS. Sur NaCl 50 mM, les organes foliaire et racinaire se chargent en cet ion pour atteindre respectivement 0,16 et 0,44 méq.g-1 MS.
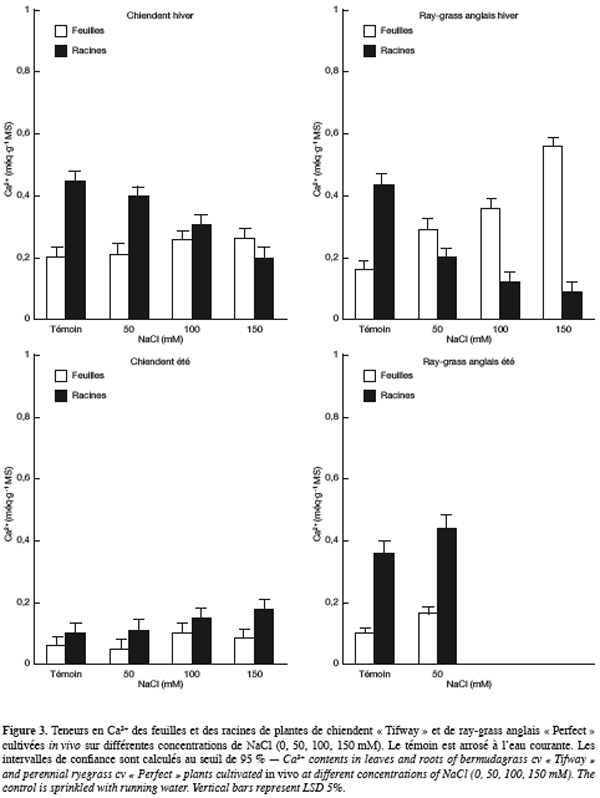
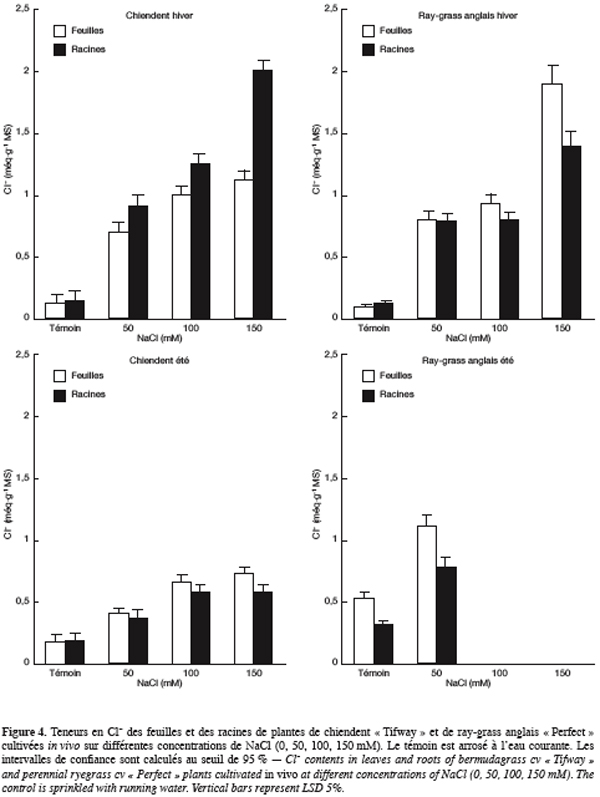
35Le chlorure. Pendant la saison hivernale, les teneurs en chlorure des feuilles et des racines de plantes de chiendent cultivées sur le milieu témoin sont faibles et comparables (0,13 méq.g-1 MS) (Figure 4). L’élévation des doses de NaCl dans l’eau d’arrosage charge clairement ces organes en Cl-, cependant, ce sont les racines qui s’en chargent le plus. Sur NaCl 150 mM, les teneurs en chlorure des feuilles et des racines arrivent respectivement à 1,12 et 2 méq.g-1 MS, soit 8 à 15 fois plus que les plantes témoins.
36Chez le ray-grass anglais, les plantes témoins présentent également les teneurs en Cl- les plus faibles et sont de 0,10 et 0,13 méq.g-1 MS, respectivement pour les feuilles et les racines. Les teneurs en cet ion augmentent avec la concentration saline et deviennent plus élevées dans les feuilles que dans les racines dès NaCl 100 mM. Sur 150 mM, elles atteignent 1,40 et 1,90 méq.g-1 MS, respectivement pour les racines et les feuilles, soit 11 à 19 fois plus que les plantes témoins.
37Pendant la saison estivale et chez le chiendent arrosé avec la solution témoin, les teneurs en Cl- des feuilles et des racines sont faibles et comparables et sont de l’ordre de 0,18 méq.g-1 MS. La salinité de la solution d’arrosage charge de plus en plus les organes de la plante en cet anion et ce sont les feuilles qui s’en chargent sensiblement plus à de fortes concentrations en NaCl. Sur 150 mM NaCl, les teneurs des feuilles et des racines en Cl- atteignent respectivement 0,73 et 0,57 méq.g-1 MS, soit 3 à 4 fois plus que les plantes témoins.
38Chez le ray-grass anglais, les teneurs en Cl- passent de 0,53 et 0,31 méq.g-1 MS sur le témoin à 1,11 et 0,77 méq.g-1 MS sur NaCl 50 mM, respecti-vement dans les feuilles et les racines.
4. Discussion
39Les teneurs en Na+ et Cl- des plantes de chiendent cultivées en période hivernale sont nettement plus élevées dans les racines que dans les feuilles. La situation s’inverse pendant la période estivale et les teneurs en ces 2 ions deviennent légèrement plus élevées dans les feuilles. Ces caractéristiques ioniques permettent de dire que les plantes de chiendent se comportent à la fois comme des « excluders » en hiver et comme des « includers » en été. Toutefois, les teneurs foliaire et racinaire en Na+ et Cl- nettement plus élevées en hiver qu’en été ne confirment pas cette hypothèse. Cette espèce étant de saison chaude et entrant en repos végétatif pendant l’hiver pourrait expliquer ce type de comportement.
40Le ray-grass anglais se comporte de la même manière en hiver et en été et accumule plus de Na+ et Cl- dans ses feuilles que dans ses racines. Il apparaît ainsi comme une espèce du type « includer » alors qu’elle est classée comme une espèce moyennement sensible à la salinité (Lemaire, 1993). Chez les 2 espèces étudiées, Cl- est de manière générale plus représenté dans les tissus foliaire et racinaire que Na+.
41Les glycophytes, plantes poussant dans les sols non salés, paraissent généralement incapables d’assurer à la fois un transfert important d’ions des racines vers les feuilles ainsi qu’une compartimentation cellulaire efficace. D’après Greenway et Munns (1980), les plantes les plus résistantes sont celles qui évitent une absorption trop importante d’ions. Certaines glycophytes, comme le cotonnier ou l’orge, transportent et accumulent de grandes quantités de Na+ dans leurs feuilles.
42Les espèces incapables de compartimenter Na+ dans leurs feuilles sont nettement plus sensibles à la salinité. En effet, ces espèces semblent peu efficaces pour abaisser la concentration cytoplasmique de Na+, ce qui est peut être l’une des causes profondes de leur sensibilité au niveau cellulaire, mais l’incapacité de débarrasser le cytoplasme de Na+ a pour conséquence que cet ion est facilement transporté dans le phloème de ces plantes (Zid, Grignon, 1986). L’exclusion continuelle de Na+ vers le bas de la plante traduit le comportement d’exclusion.
43Les teneurs en K+ des organes foliaire et racinaireø sont plus élevées chez le chiendent que chez le ray-grass anglais surtout en période estivale. Cette caractéristique ionique se rencontre généralement chez les espèces où la teneur élevée en K+ est corrélée avec la tolérance au sel (Taleisnik-Gertel, Tal, 1986). Chez les 2 espèces étudiées, la salinité du milieu a pour effet de diminuer les teneurs en K+ des organes végétatifs en période hivernale et de les augmenter, au contraire, en période estivale.
44Le rapport K+/Na+ de l’accumulation dans les racines et les feuilles de chiendent diminue avec la salinité de l’eau mais reste en faveur de K+, tant que la concentration en NaCl de l’eau d’arrosage ne dépasse pas 50 et 100 mM, respectivement dans les feuilles des plantes cultivées en hiver et dans les feuilles et les racines des plantes cultivées en été (Tableau 2). Weimberg et al. (1984) a montré que la tolérance à la salinité chez le blé serait due à une forte sélectivité en faveur de K+ témoignant de l’aptitude de cette espèce à discriminer le K+ et le Na+ au niveau de l’absorption racinaire et du transport dans les parties aériennes.
45Pour les racines des plantes de chiendent cultivées en période hivernale, le rapport K+/Na+ reste parfaitement constant et inférieur à 1 même chez les plantes témoins. Ceci signifie que Na+ est toujours prédominant, dans ces organes, relativement à K+.
46K+/Na+ diminue également dans les feuilles et les racines de ray-grass anglais avec la salinité de l’eau d’arrosage. Ce rapport est légèrement en faveur de K+ uniquement dans les racines des plantes témoins cultivées en période estivale. L’accumulation de K+ dans les feuilles des plantes témoins cultivées en hiver est égale à celle de Na+. Pour toutes les doses de NaCl, le rapport K+/Na+ est largement inférieur à 1 dans les feuilles et les racines de cette espèce. Ceci signifie que la teneur en Na+ surpasse toujours celle en K+.
47Chez le chiendent comme chez le ray-grass anglais, les teneurs en Ca2+ des feuilles et des racines sont plus faibles en été qu’en hiver. Les feuilles accumulent généralement plus de K+ que de Ca2+. La salinité de la solution d’arrosage ne modifie pas la teneur des feuilles de chiendent en ce cation, par contre, elle l’augmente nettement chez le ray-grass anglais. Les teneurs des racines en Ca2+ diminuent en période hivernale avec la concentration de NaCl chez les 2 espèces et augmentent, par contre, en période estivale.
5. Conclusion
48Les caractéristiques ioniques des plantes de chiendent, variété Tifway, diffèrent de celles du ray-grass anglais, variété Perfect, et sont dépendantes aussi de la saison de culture. En effet, le chiendent apparaît à la fois comme une plante inclusive et exclusive de Na+ et Cl- étant donné que les teneurs en ces 2 ions sont plus élevées en période hivernale qu’en période estivale et que leur accumulation est plus importante dans les racines que dans les feuilles pendant la saison hivernale.
49Le ray-grass anglais, par contre, présente un caractère inclusif vis-à-vis de Na+ et Cl- bien qu’il montre une sensibilité particulière à la salinité pendant la saison estivale ayant eu pour conséquence la mortalité de toutes les plantes soumises à des doses de NaCl au-delà de 50 mM.
50En hiver, les feuilles de chiendent restent fortement sélectives en faveur de K+ tant que la concentration de NaCl n’a pas dépassé 50 mM, en dépit de la forte prédominance de Na+ dans les racines. En été, les racines comme les feuilles deviennent très sélectives en faveur de K+ sauf en milieu fort salin (150 mM en NaCl). Le ray-grass anglais, au contraire, se caractérise par la prédominance de Na+ dans ses tissus foliaires et racinaires, ce qui peut perturber l’alimentation potassique chez cette espèce.
51En définitive, le chiendent reste plus tolérant à la salinité que le ray-grass anglais surtout en climat chaud, toutefois et en dépit de sa sensibilité à la salinité, cette dernière espèce se caractérise par un excellent comportement hivernal. Ainsi, dans les conditions tunisiennes, ces 2 espèces se complètent parfaitement et par leur association on obtient, tout au long de l’année, un tapis gazonné satisfaisant.
Bibliographie
Ehret DL., Radmann RE., Harvery BI., Cipywnyk A. (1990). Salinity induced calcium deficiencies in wheat and barley. Plant Soil 128, p. 143–151.
Greenway H., Munns R. (1980). Mechanisms of salt tolerance in non-halophytes. Ann. Rev. Plant Physiol. 31, p. 149–190.
Hajji M. (1980). La responsabilité de la racine dans la sensibilité du Laurier-rose au chlorure de sodium. Physiol. Vég. 18 (3), p. 505–515.
Hamza M. (1977). Action de différents régimes d’apport du chlorure de sodium sur la physiologie de deux légumineuses : Phaseolus vulgaris (sensible) et Hedysarum carnosum (tolérante). Relations hydriques et relations ioniques. Thèse d’état. Paris : Université de Paris VII, 252 p.
Jendoubi S. (1997). Contribution à la caractérisation physiologique et biochimique de parois racinaires chez 2 espèces de blé : Triticum durum (Ben Béchir) et Triticum aestivum (Tanit) cultivées en milieu salin. Tunis : DEA de la Faculté des Sciences de Tunis, 86 p.
Lemaire F. (1993). Qualité de l’eau d’irrigation. Paysages et actualités, novembre-décembre, p. 24–26.
Levigneron A., Lopez F., Vansuyt G., Berthomieu P., Fourcroy P., Casse-Delbart F. (1995). Les plantes face au stress salin. Cah. Agric. 4, p. 263–273.
Levitt J. (1980). Salt and ion stress. In Levitt J. (eds). Response of plant to environmental stresses. Vol II, water radiation, salt and others stresses. New York: Academic Press, p. 365–406.
Nimbalkar JD., Joshi GV. (1975). Effect of increasing salinity on germination, growth and mineral metabolism of sugarcane var. co. 740. J. Biol. Sci. 18, p. 55–63.
Piri K. (1991). Contribution à la sélection in vitro de plantes androgéniques de blé pour leur tolérance au NaCl. Thèse de Doctorat. Gembloux, Belgique : Faculté des Sciences agronomiques de Gembloux, 168 p.
Slama F. (1986). Effet du nitrate d’ammonium sur le degré de tolérance à une forte dose de NaCl de dix variétés de blé. Colloque sur les végétaux en milieu aride, Jerba (Tunisie), 8-10 septembre 1986. Tunis : Agence de coopération culturelle et technique, p. 460–473.
Smith FA. (1973). The internal control of nitrate uptake into excised barley roots with differing salt contents. New Phytol. 72, p. 769–782.
Soltani A. (1988). Analyse des effets de NaCl et de la source d’azote sur la nutrition minérale de l’orge. Thèse de Doctorat d’état. Tunis : Faculté des Sciences de Tunis, 322 p.
Taleisnik-Gertel E., Tal M. (1986). Potassium utilisation and fluxes in wild salt-tolerant relatives of the cultivated tomato. Physiol. Plant. 67, p. 415–420.
Weimberg R., Lerner HR., Poljakoff-Mayber A. (1984). Changes in growth and water-soluble solute concentrations in Sorghum bicolor stressed with sodium and potassium salts. Physiol. Plant. 62, p. 472–480.
Yakoubi-Tej M. (1992). Rôle des interactions ioniques dans la tolérance du triticale à NaCl. Tunis : Faculté des Sciences de Tunis, 63 p.
Zid E., Grignon C. (1986). Effets comparés de NaCl, KCl et Na2SO4 sur la croissance et la nutrition minérale de jeunes Citrus aurantium L. Oecol. Plant. 7 (4), p. 407–416.
Pour citer cet article
A propos de : Faouzi Haouala
Institut Supérieur agronomique. Département des Sciences horticoles. 4042 Chott Mariem, Sousse (Tunisie). E-mail : faouzi.haouala@laposte.net
A propos de : Hanen Ferjani
Institut Supérieur agronomique. Département des Sciences horticoles. 4042 Chott Mariem, Sousse (Tunisie).
A propos de : Salem Ben El Hadj
Institut National agronomique de Tunisie. 43, avenue Charles Nicolle. 1002 Tunis Belvédère (Tunisie).