


- Accueil
- volume 9 (2005)
- numéro 2
- Restoration of calcareous grasslands: the role of the soil seed bank and seed dispersal for recolonisation processes


Visualisation(s): 4569 (26 ULiège)
Téléchargement(s): 174 (2 ULiège)

Restoration of calcareous grasslands: the role of the soil seed bank and seed dispersal for recolonisation processes

Résumé
Restauration des pelouses calcicoles : le rôle de la dispersion et de la banque de graines du sol dans les processus de recolonisation. La banque de graines est un élément central pour la restauration des communautés végétales. Nous passons en revue les méthodes de mesure de la persistance des graines et les types de banques de graines. La composition floristique et la banque de graines ont été comparées dans 18 sites représentant des successions ou des plantations dans l’ouest de l’Allemagne. La richesse en espèces décroît après l’abandon de la gestion alors qu’environ 25 % des espèces survivent dans la banque de graines. Après mise à blanc, la richesse spécifique de la végétation s’accroît rapidement et la composition floristique tend vers celle d’une pelouse calcicole. Les espèces avec une banque de graines permanente ou des capacités de dispersion à longue distance présentent la plus grande probabilité de se rétablir à court terme.
Abstract
Seed bank is a central topic for plant communities restoration. We review the methods to measure seed persistence and the different seed bank types. Floristic composition and seed bank were compared among 18 sites representing various stages of succession or afforestation in West Germany. Species richness decreased in the vegetation after abandonment while about 25% of the species survived in the seed bank. After clear cutting, species richness was restored rapidly in the vegetation and floristic composition became more similar to typical calcareous grassland within five years. Species with a permanent seed bank or with the capacity of long-distance dispersal were more successful to re-establish within short terms.
Table des matières
1. Introduction
1Calcareous grasslands formerly were very common in parts of central and southern Germany. During the last century they suffered a strong decline in size and number due to the reduction of sheep raising and shepherding (Poschlod, WallisDeVries, 2002). Many pastures were abandoned, lay fallow or were afforested (WallisDeVries et al., 2002). Nowadays, nature conservationists try to preserve the remaining calcareous grasslands and also to restore sites that had been abandoned and have been overgrown by shrubs and trees. The question in this case is: Is it possible to restore them ? And would it be possible to bring back the species diversity that had been there before ?
2Referring to the fact that after several years of successional growth very often there are not many grassland species left in the actual vegetation, it is important to look for sources of recolonisation for plant species (Bakker et al., 1996a). One of these sources is the soil seed bank. Therefore, this article first summarises different types of seed banks and then provides a case study for the restoration of calcareous grassland sites in southwest Germany.
2. The soil seed bank
3Seed banks –also called diaspore banks (Poschlod et al., 2004)– consist of seeds and fruits as well as of vegetative parts of plants. In the case of mosses and ferns they also include spores. Seeds are able to remain viable in the soil for different periods of time depending on species but also soil conditions (Priestley, 1986; Poschlod et al., 2004). The longevity of a seed is decisive for how long a species is present in the seed bank after the supply of fresh seeds has been interrupted, e.g. after grassland species have been replaced by grasses or shrubs.
4How to measure persistence ? There are various ways to estimate the longevity of seeds in the soil. One method is to bury seeds in the soil and take out samples after certain periods of time as it was done for example by Duvel (1902). This burial experiment was analysed by Toole and Brown (1946). They could show that after 39 years of burial, 36 of 109 species were still alive and able to germinate. New techniques even allow to assess the age of buried seeds directly by measuring C-14 levels (McGraw et al., 1991; Moriuchi et al., 2000).
5An indirect method is to analyse the seed bank composition along successional gradients. In this case, the state of succession reflects time of after abandonment and can be related to the age of seeds in the soil (Bakker et al., 1996b).
6Similar is the analysis of the seed bank after environmental changes e.g. afforestations of calcareous grasslands. If viable seeds are found and the last floristic proof is known, the time-span of the seeds outliving in the soil can be calculated (Poschlod et al., 1998).
7Further methods are to investigate the seasonal dynamics of the seed bank (relation of seed input, germination and decay of seeds), the depth distribution of seeds in the soil and the seed size and shape as predictors for seed longevity (Poschlod et al., 2004).
2.1. Seed bank classification
8The knowledge on seed longevity leads to a classification described by Thompson (1992, 1993) where three types are defined:
9– Transient. Viable seeds remain in the soil for less than one year.
10– Short-term persistent. Seeds persist in the soil for one to five years. This type provides a buffer for low seed production in unfavourable years.
11– Long term persistent. The longevity of seeds in the soil is larger than five years. Especially this type is able to contribute to the regeneration of calcareous grasslands after management measures.
12Poschlod and Jackel (1993) created four types referring to the seasonal dynamics of seed rain/seed bank and depth distribution in soil. They used diaspore traps to catch the seed rain and soil samples to analyse the depth distribution of seeds:
13A further method to refine the seed bank classification allowing to rank species according to their persistence is the Longevity-Index (Bekker et al., 1998). It uses published data on seed longevity with species being classified as transient (t), short-term (sp) or long-term persistent (lp). The total number of records is taken to calculate:
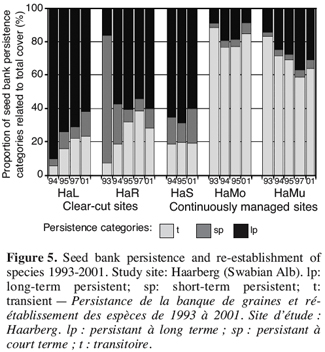
14LI = (sp + lp) / (t + sp + lp)
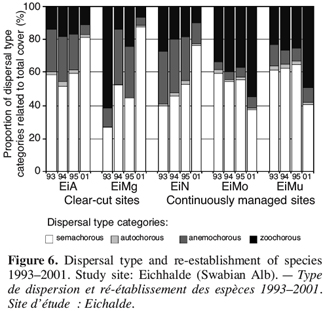
15The index ranges from 0 (strictly transient) to 1 (strictly persistent). For example, for Potentilla neumanniana the number of records is t = 4, sp = 1 and lp = 1. Thus it has a longevity-index of 0,33.
16An indicator for seed longevity is not only the depth distribution of seeds in the soil but also their mass, size and shape. Bekker et al. (1998) could prove a significant correlation between depth distribution and the combined seed attribute M x ÷Vs, where ÷Vs is the variation of seed dimensions. Briefly, the smaller a seed the higher its proportion in deeper soil layers. In a multiple regression, Bekker et al. (1998) combined this correlation with five classes of the Longevity Index. It is shown that species with a high Longevity-Index tend to be small and well represented in deeper soil layers.
2.2. The importance of the seed bank for calcareous grassland restoration
17For conservation issues, the seed bank is important to preserve species of the local species pool – about 1/4 to 1/3 of the species of calcareous grasslands build up a long-term persistent seed bank. If the seed density in the soil is sufficient, this potential can be readily activated when the environmental conditions are being restored with the help of management measures. However, without a proper and ongoing management, the persistent seed bank can be depleted when seeds are germinating but are not able to establish themselves as plants.
3. A case study: Re-establishment of calcareous grasslands after abandonment or afforestation by clear-cutting
18This case study has been carried out on 18 sites, situated in the Neckar Basin and on the Swabian Alb in south-western Germany (Kiefer, 1998; von Blanckenhagen, 2002).
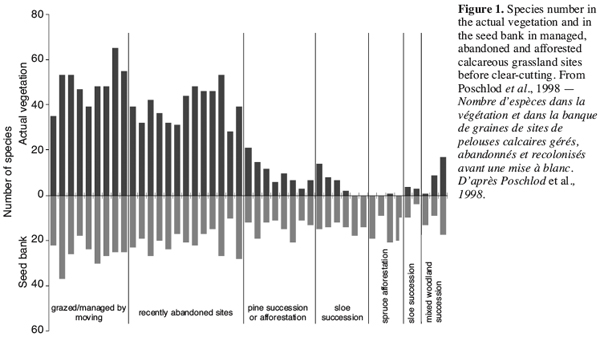
19The sites included sloe-, pine- and robinia-successions, spruce or pine afforestations or mixed woodland successions. The age of the successional states or afforestations respectively ranged from 5 to 55 years leading to different floristic compositions of the remnant vegetation in the herbal layer. In many cases, herbal species had disappeared completely underneath the canopy of shrubs and trees. Figure 1 points out that species numbers in the actual vegetation decrease after abandonment or afforestation and that about a quarter of the species is able to persist in the seed bank in some cases for more than 50 years (original data in Kiefer, 1997).
20The vegetation development of the sites was investigated using five permanent plots (4 m2 each) from the point of time before clear-cutting in 1990/1991 or 1992/1993 until 2001. A general trend of the vegetation development of the clear-cut plots was a rapid increase of species numbers within the first three years. Due to the occurrence of species from different sociological groups, the total number of species of the clear-cut sites was frequently higher than the one of the calcareous grassland controls.
21Especially species of the Stellarietea and Artemisietea, species adapted to high levels of disturbance, prospered well on the open soils in the first years, but receded strongly again until 2001. Festuco-Brometea-species were steadily increasing. However, in comparison of all sites, they were still significantly less abundant on the clear-cut sites than on the continuously managed sites at the end of the study period. By that time, shrub species (mainly Prunus spinosa and Cornus sanguineus) still exhibited up to 10% vegetation cover, which points out the necessity of ongoing management.
22In the following, one of the study-sites will be shown in more detail: The “Haarberg” on the edge of the Swabian Alb. It had been afforested with spruce in the 1960’s and was finally clear-cut in 1992. The above mentioned trends can be seen in figure 2.
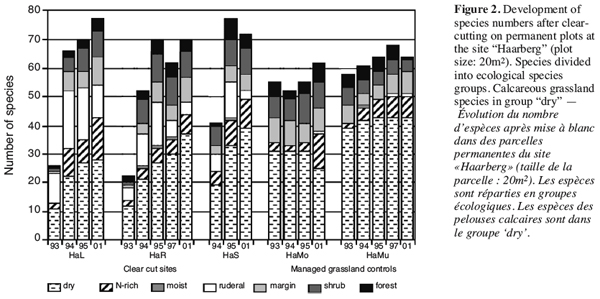
23After clearing the site, there had been no species left in the herbal layer. In the first years, ruderal species like Cirsium arvense, Daucus carota and Melilotus officinalis reached up to 20–30% cover. Their cover finally decreased to less than 5% in 2001. On the other hand, calcareous grassland species were continuously increasing over the years. Among these, Euphorbia cyparissias and Carex flacca were abundant from the start.
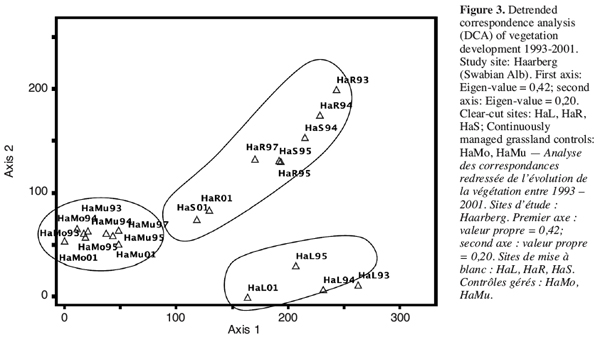
24A detrended correspondence analysis (Figure 3) clarifies the approach of the plots that had been clear-cut to the continuously managed plots; their floristic compositions become more and more similar.
4. Biological traits and re-establishment of plant species
25The re-establishment of species was furthermore analysed referring to several biological traits, which included generative as well as vegetative traits. Different data sets on information on plant functional types including own investigations were used (Poschlod et al., 2000). In the following, we relate some of the analysed traits to the vegetation development of selected sites (Eselrain–spruce afforestation beginning of the 1970’s, clear-cut in autumn 1993; Haarberg–spruce afforestation in the 1960’s, clear-cut in winter 1992/1993; Eichhalde–abandonment in the 1960’s, clear-cut of a sloe successional stage in winter 1991/1992): the results concerning seed production, seed mass, seed bank persistence and dispersal potential are briefly presented here.
26Seed production is an important trait for the persistence at the site as well as for dispersal, especially within short distances. A large seed production enhances the chance to find safe-sites (Jakobsson, Eriksson, 2000). However, this does not ensure germination and growth in any kind of vegetation type. Seedlings from large seeds have better chances to survive, especially in dense vegetation, due to their nutrient resources. There is a trade-off between seed size and number that has been described by Leishman (2001).
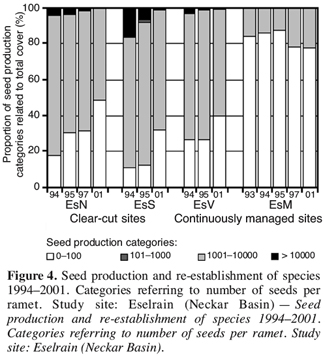
27In the first analysis, seed mass was found to have a low or no impact on recolonisation processes in the analysed grasslands. Species with a high seed production, on the other hand (i.e. more than 100 seeds per ramet, e.g. Mycelis muralis), were significantly more abundant on the clear-cut sites than on the continuously managed sites (controls), referring to species numbers and total cover. This is shown at the example of the study site “Eselrain” (Figure 4). They benefited from the low cover of the herbal layer after clear-cutting.
28The longevity of the seed bank is an important aspect in restoration ecology. The ability to build up a permanent seed bank is a means for plants to “disperse in time” and to survive unfavourable conditions. Germinable seeds in the soil are part of the population of a species even if it has already disappeared from the actual vegetation (Poschlod, 1991). In the case study, the importance of the seed bank for the restoration of calcareous grasslands could be shown (Figure 5). Long-term persistent species are “the first to reappear” (e.g. Euphorbia cyparissias, Carex ornithopoda). On the clear-cut sites, the number of species and the total cover of species with a persistent seed bank was significantly higher than on the control-plots, which is shown at the example of the study site “Haarberg”. These species had better chances to establish from seeds if after clear-cutting the remnant herbal layer offered a number of gaps.
29Species without persistent seed banks rely on the ability to disperse in space. With the presence of bare ground after clear-cutting, anemochorous plants may be successful colonisers. In this study, in the first three years, numbers of species and total cover of species with wind-dispersed seeds were significantly higher on the clear-cut sites than on the grassland controls (e.g. Cirsium arvense, Picris hieracioides; Figure 6).
30If recolonisation processes occur, they strongly depend on the local species-pool (Eriksson, 1993; Zobel et al., 1997). Kiefer, Poschlod (1996) could show that short-distance dispersal is of greatest importance concerning the recolonisation of restored grassland sites and that it has a strong influence on recolonisation patterns. Seeds with morphological adaptations for long-distance dispersal (e.g. zoochorous: hooks, anemochorous: parachutes) might be transported over long distances although the majority of them often remains close to the mother plant (Willson, 1993). For calcareous grasslands, sheep were shown to be very important vectors to transport seeds more than just a few metres (Fischer et al., 1996, Poschlod et al., 1998). Despite the fact that some of the clear-cut sites were grazed afterwards, our data set did not present the role of the sheep as clearly as we expected. This might also depend on the analysis where we used the morphological classification after Luftensteiner (1982) instead of the real dispersal potential by sheep of each species, what would be by far more realistic (Poschlod et al., 2004). This data will be only available in the near future.
5. Conclusion
31In this study it could be shown that calcareous grasslands can be restored after clear-cutting if a continuous management (grazing/mowing) of the sites is guaranteed. But even after nine or ten years of management there are still differences in species composition between clear-cut sites and controls.
32Recolonisation processes follow certain patterns where species with a permanent seed bank or with the capacity of long-distance dispersal are more successful to re-establish within short terms. Conclusively, dispersal in time as well as in space is crucial for the recreation and conservation of calcareous grassland communities.
Bibliographie
Bakker JP., Poschlod P., Strykstra RJ., Bekker RM., Thompson K. (1996a). Seed banks and seed dispersal: important topics in restoration ecology. Acta Bot. Neerl. 45, p. 461–490.
Bakker JP., Bakker ES., Rosén E., Verweij GL., Bekker RM. (1996b). Soil seed bank composition along a gradient from dry alvar grassland to Juniperus shrubland. J. Veget. Sci. 7, p. 165–176.
Bekker RM., Bakker JP., Grandin U., Kalamees R., Milberg P., Poschlod P., Thompson K., Willems JH. (1998). Seed size, shape and vertical distribution in the soil: indicators of seed longevity. Funct. Ecol. 12, p. 834–842.
Duvel JWT. (1902). Seeds buried in soil. Science 17, p. 872–873.
Eriksson O. (1993). The species-pool hypothesis and plant community diversity. Oikos 68, p. 371–374.
Fischer SF., Poschlod P., Beinlich B. (1996). Experimental studies on the dispersal of plants and animals on sheep in calcareous grasslands. J. Appl. Ecol. 33, p. 1206–1222.
Jakobsson A., Eriksson O. (2000). A comparative study of seed number, seed size, seedling size and recruitment in grassland plants. Oikos 88, p. 494–502.
Kiefer S., Poschlod P. (1996). Restoration of fallow or afforested calcareous grasslands by clear-cutting. In Settele J., Margules CR., Poschlod P., Henle K. (Eds.) Species survival in fragmented landscapes. Dordrecht, The Netherlands: Kluwer Academic Publishers, p. 209–218.
Kiefer S. (1998). Untersuchungen zur Wiederherstellung brachgefallener oder aufgeforsteter Kalkmagerrasen. Berichte des Institutes für Landschafts–und Pflanzenökologie der Universität Hohenheim; Beiheft 7. Ostfildern, Germany: Verlag Günter Heimbach, Dissertation. 309 S.
Leishman MR. (2001). Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality. Oikos 93, p. 294–302.
Luftensteiner HW. (1982). Untersuchungen zur Verbreitungsbiologie von Pflanzengemeinschaften an vier Standorten in Niederösterreich. Bibl. Bot. 135, p. 1–68.
McGraw JB., Vavrek MC., Bennington CC. (1991). Ecological genetic variation in seed banks. I. Establishment of a time-transect. J. Ecol. 79, p. 617–626.
Moriuchi KS., Venable DL., Pake CE., Lange T. (2000). Direct measurement of the seed bank age structure of a Sonoran Desert annual plant. Ecology 81, p. 1133–1138.
Poschlod P. (1991). Diasporenbanken in Böden – Grundlagen und Bedeutung. In Schmid B., Stöcklin J. (Hrsg.) Populationsbiologie der Pflanzen. Basel, Switzerland: Birkhäuser Verlag, p. 15–35.
Poschlod P., Jackel AK. (1993). The dynamics of the generative diaspore bank of calcareous grassland plants. I. Seasonal dynamics of diaspore rain and diaspore bank in two calcareous grassland site of Suebian. Alb. Flora 188, p. 49–71.
Poschlod P., WallisDeVries M. (2002). The historical and socioeconomic perspective of calcareous grasslands – lessons from the distant and recent past. Biol. Conserv. 104, p. 361–376.
Poschlod P., Kiefer S., Tränkle U., Fischer S., Bonn S. (1998). Plant species richness in calcareous grasslands as affected by dispersability in space and time. Appl. Veget. Sci. 1, p. 75–90.
Poschlod P., Kleyer M., Tackenberg O. (2000). Databases on life history traits as a tool for risk assessment in plant species. Z. Ökologie Nat. 9, p. 3–18.
Poschlod P., Tackenberg O., Bonn S. (2004). Plant dispersal potential and its relation to species frequency and coexistence. In van der Maarel E. (Ed.). Vegetation Ecology. London: Blackwell, in press.
Priestley DA. (1986). Seed Aging. Ithaca: Cornell University Press.
Thompson K., Baster K. (1992). Establishment from seed of selected Umbelliferae in unmanaged grassland. Funct. Ecol. 6, p. 346–352.
Thompson K., Band SR., Hodgson JG. (1993). Seed size and shape predict persistence in soil. Funct. Ecol. 7, p. 236–241.
Toole EH., Brown E. (1946). Final results of the Duvel buried seed experiment. J. Agric. Res. 72, p. 201–210.
von Blanckenhagen B. (2002). Funktionale Analyse der Wiederbesiedlung von Kalkmagerrasenstandorten nach Rodung. [Functional analysis of the recolonisation of calcareous grassland sites after clear-cutting] – Philipps — Universität Marburg, Fachbereich Biologie, Fachgebiet Naturschutz II, Master thesis.
WallisDeVries M., Poschlod P., Willems J. (2002). Challenges for the conservation of calcareous grasslands in northwestern Europe: integrating the requirements of flora and fauna. Biol. Conserv. 104, p. 265–273.
Willson MF. (1993). Dispersal mode, seed shadows and colonization patterns. Vegetatio 107/108, p. 261-280.
Zobel M. (1997). The relative role of species pools in determining plant species richness: an alternative explanation of species coexistence? Trends Ecol. Evol. 12, p. 266–269.