


- Accueil
- volume 9 (2005)
- Improvement of lupin seed valorisation by the pig with the addition of a-galactosidase in the feed and the choice of a suited variety


Visualisation(s): 3476 (48 ULiège)
Téléchargement(s): 106 (2 ULiège)

Improvement of lupin seed valorisation by the pig with the addition of a-galactosidase in the feed and the choice of a suited variety

Notes de la rédaction
Received 10 April 2005, accepted 29 August 2005.
Résumé
Amélioration de la valorisation du lupin par le porc grâce à l’adjonction d’a-galactosidase à l’alimentation et au choix d’une variété adaptée. Trois expériences ont été menées pour estimer si un supplément d’a-galactosidase améliorait l’utilisation des graines de lupin par les porcs en croissance – engraissement et pour déterminer la meilleure variété de lupin en présence d’a-galactosidase. Dans l’expérience 1, trois régimes iso-azotés (iso-N) et iso-énergie nette (iso-NE) contenant 15 % de tourteau de soja (régime SBM) ou 20 % de graines de lupin (Lupinus albus, var. Arès), en présence ou non d’un supplément d’a-galactosidase (régimes ‘lupin’ et ‘lupin + a-gal’, respectivement), ont été ingérés par 12 porcs munis d’une canule caecale, selon un dispositif expérimental complètement aléatoire. La digestibilité iléale des a-galactosides tendait à augmenter avec le régime lupin + a-gal. La rétention azotée des porcs mangeant le régime tourteau de soja était similaire à celle des animaux nourris avec le régime lupin + a-gal et supérieure à celle observée chez les animaux recevant le régime lupin. Les résultats suggéraient que les a-galactosides étaient les principaux facteurs antinutritionnels dans la graine de lupin pour le porc. Les mêmes régimes ont été étudiés dans un essai de production (expérience 2) impliquant 48 porcs dont le poids vif variait de 30 à 105 kg. La durée d’engraissement était similaire pour les régimes tourteau de soja et lupin + a-gal (107 j), et plus longue pour le régime lupin (123 j). Avec le régime lupin, nous avons observé un plus faible gain quotidien moyen (ADG, -16 %) et un indice de consommation supérieur (FCR, +8 %) que pour les deux autres régimes. Le lard dorsal des animaux nourris avec le régime lupin contenait moins d’acides gras polyinsaturés (PUFA) et avait un rapport w6/w3 inférieur. Dans l’expérience 3, trois régimes iso-N et iso-NE contenant différentes variétés de lupin (Lupinus albus cv Lublanc et Amiga ; Lupinus angustifolius cv Boltensia) et un supplément d’a-galactosidase ont été étudiés sur 12 porcs munis d’une canule au caecum, comme dans l’expérience 1. Les résultats montraient que l’apport de protéines digestibles par le régime Boltensia était supérieur aux autres. Cependant, la variété Boltensia occasionnait une plus forte excrétion d’N urinaire que la variété Lublanc. Conséquemment, l’utilisation des protéines ingérées et digérées était assez élevée avec le régime Lublanc, intermédiaire avec le régime Amiga et faible avec le régime Boltensia. Les résultats suggéraient que la variété Lublanc était la mieux adaptée pour l’alimentation porcine lorsque celle-ci contenait un supplément d’a-galactosidase.
Abstract
Three experiments were conducted to assess whether an a-galactosidase supplement improved the utilisation of lupin seeds by growing and finishing pigs and to determine the best lupin variety in presence of a-galactosidase. In experiment 1, three iso-nitrogen (iso-N) and iso-net energy (iso-NE) diets containing 15% of soybean meal (SBM diet) or 20% of lupin seeds (Lupinus albus var. Arès), with or without an a-galactosidase supplement (‘lupin’ and ‘lupin + a-gal’ diets, respectively), were ingested by 12 pigs fitted with a caecal canula, following a completely randomized experimental design. The ileal digestibility of a-galactosides tended to increase with the lupin + a-gal diet. Nitrogen retention of pigs eating the SBM diet was similar to that of those fed with the lupin + a-gal diet and higher than that observed for pigs given the lupin diet. The results suggested that a-galactosides are the main anti-nutritional factors in lupin seeds for pigs. The same diets were investigated in a production trial (experiment 2) involving 48 pigs with a live weight of between 30 and 105 kg. The fattening period was similar for the SBM and lupin + a-gal diets (107 d) and longer for the lupin diet (123 d). With the lupin diet there was a lower average daily gain (ADG, -16%) and a higher food conversion ratio (FCR, + 8%) than with the two other diets. The backfat of pigs fed with lupin-based diets contained fewer polyunsaturated fatty acids (PUFA) and had a lower w6/w3 ratio. In experiment 3, three iso-N and iso-NE diets containing different varieties of lupin (Lupinus albus cv Lublanc and Amiga; Lupinus angustifolius cv Boltensia) and an a-galactosidase supplement were investigated in 12 pigs fitted with a caecal canula, as in Exp. 1. The results showed that the digestible protein supply by the Boltensia diet was higher than in the others. However, the Boltensia variety led to a higher urinary N excretion than the Lublanc variety. The results obtained with Amiga were intermediate. Consequently, the utilisation of ingested and digested proteins was relatively high with the Lublanc diet, intermediate with the Amiga diet and low with the Boltensia diet. The results suggested that the Lublanc variety was better adapted for pig feed that contained an a-galactosidase supplement.
Table des matières
1. Introduction
1Measures taken in accordance with the Common Agricultural Policy (CAP) of the European Union (EU) have adversely affected protein-rich plant for cereal production. In 2003, the EU (25 countries) produced between 5 and 6 Mt of plant proteins (excluding fresh forage), but the amount that was used in animal feed was higher than 25 Mt equivalent protein (UNIP, 2004). The deficiency in plant protein production in the EU therefore exceeds 75%, which means that large imports are needed, notably of soybean meal (SBM, 30.5 Mt), to ensure well-balanced feed for livestock.
2Lupin contains more proteins (36% DM) than pea (24% DM) or faba bean (28% DM) and is therefore more suitable for replacing SBM in EU livestock feed. It is also a good main crop in crop rotation, enhancing biodiversity and, like all leguminous crops, not requiring N fertilizers and having a low energy cost. Low-alkaloid varieties have already been selected, but all lupin varieties contain a-galactosides such as raffinose, stachiose and verbascose (Veldman et al., 1993) that are not digested in the small intestine of pigs due to the lack of a-galactosidase in the digestive secretions (Carré et al., 1985) and are therefore likely to interfere with nutrient absorption (Gdala et al., 1997) and to induce flatulence problems due to their fermentation in the colon (Delzenne, Robertfroid, 1994). Our research addressed the problems related to a-galactosides for growing and finishing pigs.
3The first trial investigated the effects of a partial substitution of SBM by white lupin seeds (Lupinus albus var. Arès), with or without an a-galactosidase supplement, on the ileal and faecal apparent digestibility of nutrients and on nitrogen (N) retention of animals. The same diets were later used in a production trial involving pigs with a live weight of between 30 and 105 kg and whose zootechnical performance and carcass quality were monitored. A third trial sought to compare the nutritional value of three lupin varieties (Lupinus albus var. Lublanc and Amiga, and Lupinus angustifolius var. Boltensia), that produce high yields in EU countries, when the diets contain an a-galactosidase supplement.
2. Material and methods
2.1. Experiment 1
4Experimental scheme. Twelve Pietrain ¥ Landrace castrated pigs (37.4±1.16 kg), fitted with a caecal canula and kept in digestibility cages, were put into three homogenous groups, each receiving a different diet (Table 1), following a completely randomized experimental design. The wheat-based diets differed in the protein source (‘SBM diet’ or ‘lupin diet’) and in the presence or not of an a-galactosidase supplement (‘lupin’ or ‘lupin + a-gal’). The diets were iso-N and iso-net energy (NE) and had a similar nutritional value according to the Dutch system (CVB, 1999). The diets were mixed with water (1/1 v/v) and distributed in two equal meals, at 08h00 and 20h00, at a daily intake level of 90 g kg-0.75 d-1 DM. The a-galactosidase (EC 3.2.1.22 – Roche Vitamins NV, Deinze, Belgium – Alpha-Gal 1000 L), provided in a liquid form, was diluted in water (40 g of 1000 GALU* g-1 in 49,960 L of water, equivalent to 800 GALU kg-1 of feed) and conserved in a cold room (4°C) before being mixed into the feed of pigs receiving the lupin + a-gal diet.
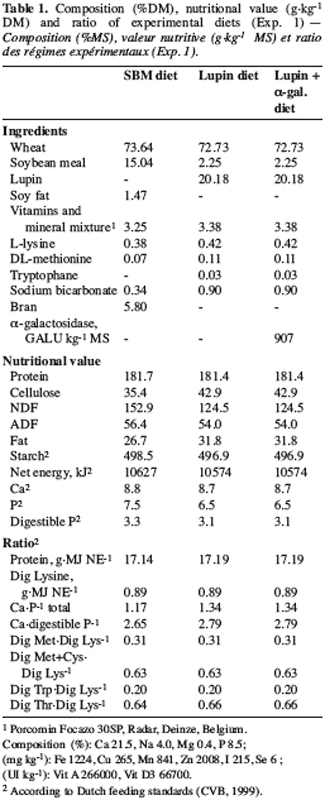
5The experiment involved seven days of animal adaptation to the feed, followed by nine days of urine and faeces collection and two days for intestinal content sampling. All urine was collected, filtrated and weighed every morning for each animal before sampling (100 mL) and freezing. Individual faeces were mixed daily, sampled and freeze-dried. The intestinal contents were collected continuously from 08h00 to 20h00. Three days before the beginning of sampling and throughout the experiment, chromic sesquioxyde (1.5 g per meal) was incorporated into the feeds to determine the nutrient flows in the gastrointestinal tract.
6Laboratory analyses. The feeds were ground (1 mm screen) before analysis of DM, ash, cellulose, fat (AOAC, 1990), protein (Dumas method, AFNOR V18-120, 1997), NDF and ADF (Van Soest et al., 1991), a-galactosides (Gdala et al., 1997) and crude energy.
7After thawing and homogenization, total N (Dumas method, AFNOR V18-120, 1997) was measured in the urine samples. The faeces were ground (1 mm) before determining DM, ash, protein (Dumas method, AFNOR V18-120, 1997) and chromic sesquioxyde (François et al., 1978). NDF, ADF (Van Soest et al., 1991), cellulose (AOAC, 1990), a-galactosides (Gdala et al., 1997) and crude energy were measured in a mixed sample of faeces excreted by each pig during the first nine days of sampling.
8After freeze-drying and grinding (1 mm), the intestinal contents were gathered from each animal and analysed for their DM, ash, cellulose (AOAC, 1990), protein (Dumas method, AFNOR V18-120, 1997), chromic sesquioxyde (François et al., 1978), crude energy and a-galactosides (Gdala et al., 1997) content.
9Statistics. The statistical analyses followed the GLM procedure in Minitab software (2000). Nitrogen retention and faecal apparent digestibility of DM, OM and N data were analysed using a two-way ANOVA (diet and day). Ileal digestibility of nutrients and faecal digestibility of fibres (NDF, ADF, cellulose), fat, crude energy and a-galactosides – measured after gathering samples from each animal – were analysed using one-way ANOVA (diet). Least square means were compared using the Tukey test (Minitab).
10The degradation of nutrients in the large intestine was calculated according to the difference in the caecal and faecal flows.
2.2. Experiment 2
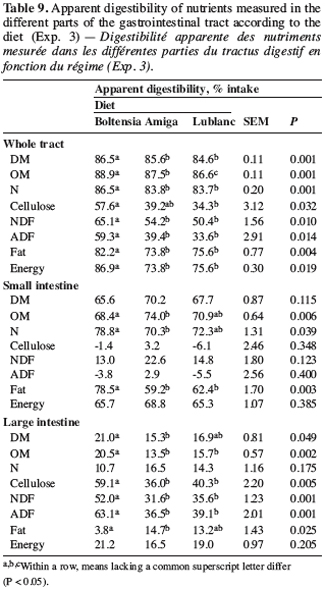
11Experimental scheme. Six groups of eight Pietrain Landrace pigs (32.3 ± 3.3 kg) were fed with the same diets used in experiment 1 (two groups of pigs per diet) during the growing-finishing period. Each group of animals contained four females and four castrates. Sawdust was used as litter and the pigs were weighed at 22, 43, 64, 78, 89, 96, 103, 110, 117, 124, 131 and 138 days. They completed the trial when they reached a live weight of 105 kg. Average daily gain (ADG) was calculated throughout the growing-finishing period. The amount of feed distributed was monitored to calculate the feed conversion ratio (FCR) per group. The pigs were slaughtered at a live weight of between 105 and 115 kg and the main traits of carcass quality were determined on 36 pigs (three females and three castrates per group).
12Laboratory analysis. The carcass quality indicators used were: carcass yield, pH and conductivity (PQM-I/Kombi, Intek GmbH, Airchach, Germany), DM (AFNOR 2001, NF V 04-401), total (AFNOR 2001, NF V 04-402) and free lipids percentage (AFNOR 2001, NF V 04-403) of the meat (Longissimus dorsi) and its fatty acid profile (Folch et al., 1957; Collomb, Buhler, 2000; ISO/IDF, 2002). A complete analysis of the nutritional value of the feeds was conducted as in experiment 1.
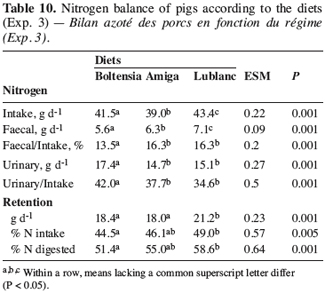
13Statistics. The statistical analyses followed the GLM procedure in Minitab software (2000). Live weight, ADG and quality parameters were analysed using a three-way ANOVA (diet, sex and group). Data on daily intake and FCR were analysed using a one-way ANOVA (diet). Least square means were compared using the Tukey test (Minitab).
2.3. Experiment 3
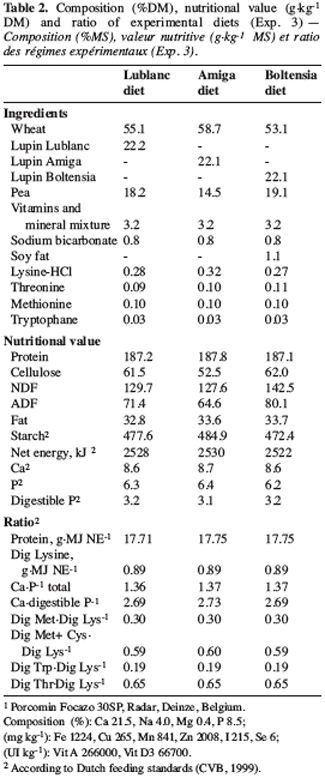
14This experiment was conducted with 12 Pietrain ¥ Landrace castrated pigs (38.1 ± 3.1 kg) fitted with a caecal canula. The experimental design was similar to that used in experiment 1. The diets were iso-N and iso-NE (Table 2) and differed according to the lupin variety used (Lupinus albus cv Lublanc and Amiga, and Lupinus angustifolius cv Boltensia). All the diets contained a supplement of a-galactosidase (800 GALU.kg-1 of feed). Sampling and analyses were conducted as in experiment 1.
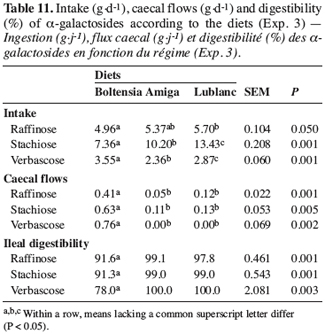
3. Results and discussion
15All the procedures used involving the animals were approved by the Comité d’éthique en expérimentation animale (protocol CRA 01/08 and CRA 03/11).
3.1. Experiment 1
16Digestibility parameters. On the whole digestive tract, most nutrients tended to be better digested with the SBM diet than with the lupin-based diets (Table 3). According to Salgado et al. (2002), the high content of undigestible NDF in lupin seeds could lead to lower faecal digestibility of DM, OM and energy. The addition of a-galactosidase to the lupin diet caused an additional decline in faecal N digestibility. A separate analysis of digestibility in the small intestine and in the large intestine is required to produce a hypothesis.
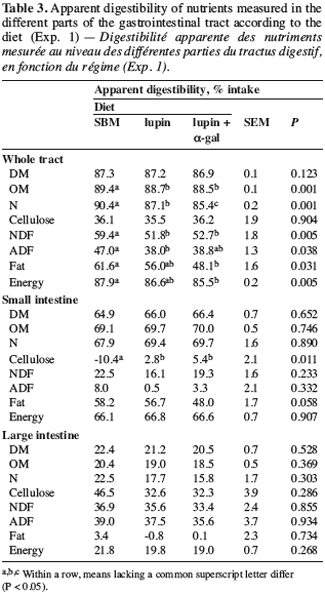
17Ileal apparent digestibility of nutrients was not influenced significantly by the diets. Apart from DM and OM, these results agree with those of Salgado et al. (2002) but not with those of Gdala et al. (1997), which showed that the addition of a-galactosidase is likely to increase DM, OM and most amino acids (AA) digestibility. In agreement with Wiggins (1984), these authors considered that the presence of a-galactosides led to a higher osmolarity of the intestinal contents, affecting the absorption of some nutrients.
18Apart from N, there was no difference in the fermentation of nutrients in the diets. According to Salgado et al. (2002), lupin fibres are partially fermented in the large intestine, which could stimulate the bacterial growth and thereby cause a higher loss of faecal N in a microbial form. Despite the addition of wheat bran to the SBM diet to balance the fibre supply, we observed a lower N digestibility in the large intestine with lupin-based diets, which probably reflected the higher microbial activity in these diets. However, lupin seeds are rich in soluble fibres that are easily fermentable (Bach Knudsen, Gonzalez, 2004) and supply a different type of fibre than bran. The negative digestibility of fat measured with lupin-based diets could be explained by a neo-synthesis of bacterial origin (Noblet, Legoff, 2001).
19Nitrogen balance. The diets did not influence the N intake (Table 4). Faecal N excretion was higher with lupin-based diets, particularly with the lupin + a-gal diet. The lupin diet led to a higher urinary N excretion than the lupin + a-gal diet; the value measured for the SBM diet was intermediate. The association of lupin with a-galactosidase reduced urinary N loss by 2.1 g.d-1, corresponding to 12.8% compared with the lupin diet, but it also resulted in a higher faecal N excretion. This observation tends to confirm the likelihood of bacterial activity being stimulated in the large intestine of pigs receiving the supplement of a-galactosidase.
20The N retention (g.d-1) of pigs fed with the lupin diet was lower than that of pigs receiving the SBM diet (Table 4). An incorporation level of 20% of white lupin seeds in growing pigs’ diets was therefore too high if their zootechnical performance was not to be affected. This effect probably derived from the a-galactosides content of the lupin seeds because the addition of a-galactosidase to this diet resulted in a similar N retention to that in pigs receiving SBM. The use of intake proteins (N retention related to N intake) and digested proteins (N retention related to digested N) confirmed the greater added value of lupin seeds in the presence of the enzyme supplement.
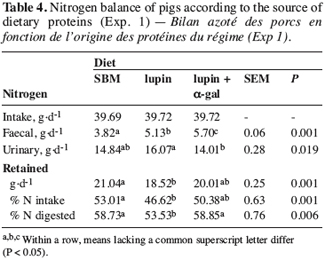
21The digestion of a-galactosides. The lupin-based diets contained more stachiose and verbascose than the SBM diet, but the raffinose content was similar (Table 5). The ileal digestibility of a-galactosides was similar for the SBM and lupin diets and tended to increase with the lupin + a-gal diet. This reflected the good activity of the enzyme supplement under our experimental conditions. In all the diets, a large part of a-galactosides was digested at the end of the small intestine, while the pigs did not produce a-galactosidase (Carré et al., 1985). This observation could indicate the presence of an active microflora in the ileum. More work is needed to investigate the influence of this microflora on the nutritional value of nutrients, the presence of bacteria often seen as being limited to the large intestine. The digestibility of raffinose and stachiose was complete at the end of the digestive tract, but the verbascose was still present in very small amounts in the faeces. Despite a significant ingestion of a-galactosides, the pigs receiving the lupin diet had no digestive problems.
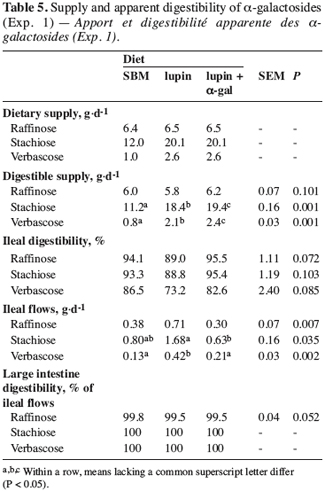
3.2. Experiment 2
22Growth performances. In all the diets, the weight of the pigs at the beginning and at the end of the trial was similar (Table 6). However, the fattening period increased significantly with the lupin diet compared with the SBM and lupin + a-gal diets. The ADG for the whole trial was lower with the lupin diet. This could stem partly from the lower ingestion of this diet, but the FCR was also higher in this diet than in the other two diets. This suggests that lupin was poorly used when it is not associated with a-galactosidase. The live weight evolution showed that the harmful effect of a-galactosides occurred mainly during the first part of the animal’s growth, up to a live weight of 55 kg, and that the weight difference persisted afterwards. The initial and final weights of the pigs, the fattening period and the ADG were not influenced significantly by animal gender (Table 6).
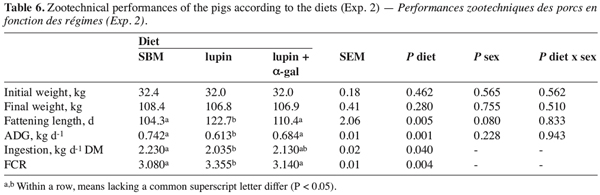
23Carcass quality. Slaughter and carcass quality parameters were not influenced by diet (Table 7) or gender (P > 0.05, apart from the pH at 24h, which was lower by 0.107 units for castrates). The absence of a diet effect could be explained by their similar nutritional value. Roth-Maier et al. (2004) showed previously that carcass characteristics were not affected by lupin inclusion (20% DM) or by lupin variety used for pig feed.
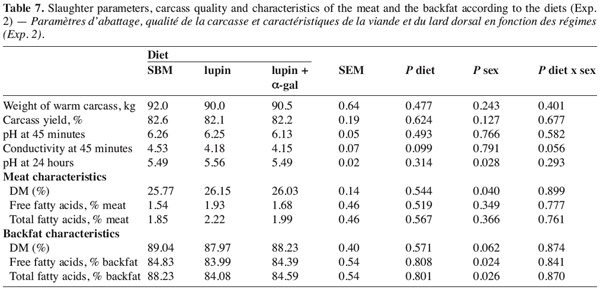
24Apart from the w6/w3 ratio, the diet did not influence the fatty acid profile of the meat. The backfat of the pigs receiving the lupin-based diets contained more monounsaturated fatty acids (MUFA) and fewer polyunsaturated fatty acids (PUFA) (Table 8) than pigs given the SBM diet. As suggested by Mourot (2001), this modification reflected the composition of dietary fat; the fat of lupin seeds being characterized by a high concentration of C18:1, while the soy fat used in the control diet was rich in C18:2. According to Warnants et al. (2001), the stability of the fatty acid profile of the meat could be explain by the fact that PUFA are more easily incorporated in lumps of fat, as backfat, than in intra-muscular fat.
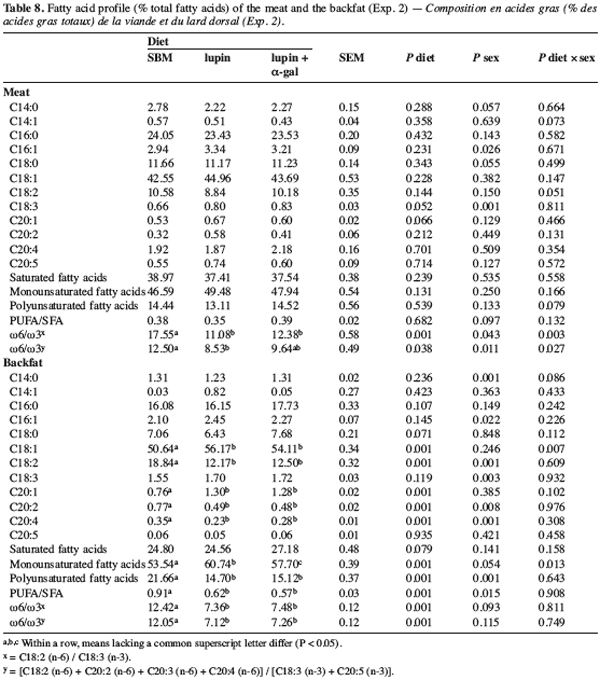
25For humans, a healthy diet requires limiting the ingestion of saturated fatty acids (SFA) in favour of PUFA. However, it is not always possible to set up an optimal fatty acid profile for human health and for technological requirements. According to Warnants et al. (2001), the optimal PUFA/SFA ratio for backfat transformation (sausages, etc.) should be 0.6-0.7. Such values were observed for pigs receiving the lupin-based diets but not for those receiving the SBM diet (0.91). From a technical point of view, such a high ratio indicates significant risks of fat oxidation and rancidness. Girard et al. (1988) considered that the C18:2 of backfat should not exceed 12–15% of total fatty acids to ensure good transformation and conservation. Lebret and Mourot (1998) recommended distributing a rich SFA diet to finishing animals in order to reduce the C18:2 content of backfat.
26The w6/w3 ratio was higher in the intra-muscular fat than in the backfat of pigs fed with the SBM diet (Table 8). This reflects a high content of C18:2 (n-3) in soy fat compared with lupin fat. Human food is also rich in w6 and the use of lupin seeds in pig feed instead of soy protein could therefore be interesting.
3.3. Experiment 3
27Digestibility parameters. The intestinal digestibility of N and fat was lower for the white lupin-based diets than for the Boltensia diet (Table 9). As suggested by Gdala et al. (1999), protein digestibility should vary greatly among cultivars, especially for blue lupins.
28In the large intestine, the fibres supplied by the Boltensia diet were better degraded than the fibres from the white lupin-based diets. This explained the differences observed in fibre digestibility in the whole tract. Blue lupins contain more fibres than white lupins because of a higher proportion of hulls, indicated by a lower 1000-seed-weight (236.0 and 247.3 g for Lublanc and Amiga, respectively, and 115.5 g for Boltensia). A large variability among cultivars in terms of fibre composition is noted in the literature (Gdala et al., 1996). Blue lupins differ from white lupins in their richness in non-cellulosic polysaccharides (Bach Knudsen, Gonzalez, 2004) and in cellulose (Cherrière et al., 2003). These characteristics could explain the better degradability in the large intestine of fibres supplied by the Boltensia diet.
29Nitrogen balance. The N intake varied slightly among the diets (P < 0.05) due to variations in feed composition (Table 10). Faecal N excretion also differed among the diets. However, the part of the N intake excreted in the faeces in both white lupin-based diets was similar and higher than that in the Boltensia diet. This observation confirmed that proteins supplied by blue lupins were better digested than those of white lupins.
30Unlike the faecal N flows, urinary N excretion was higher with the Boltensia diet than with the Amiga and Lublanc diets. The variation in N intake does not explain this difference, and the reason why digestible N in the Boltensia diet was less used remains unclear. This observation contradicts the results obtained by Bach Knudsen and Gonzalez (2004), which showed that, in rats, the biological value and the net utilisation of proteins were higher for blue lupins than for white lupins. However, our results are in agreement with the study of Rahman et al. (1997) mentioning unexplained disturbances in N metabolism and a low retention value of the absorbed N with diets containing blue lupins in rats.
31Finally, the N retention (g.d-1) was lower with the Amiga and Boltensia diets than with the Lublanc diet (Table 10). This also contradicts the findings of several studies, suggesting that white lupins do not lead to good zootechnical performance in growing-finishing pigs, compared with blue lupins (Gdala et al., 1996, Dunshea et al., 2001). However, none of these studies compared both type of lupins in the presence of an a-galactosidase supplement. The reasons given by the authors also tend to vary and focus on the variety characteristics related to the composition and the physico-chemical properties of fibres (Dunshea et al., 2001), the presence of a-galactosides (Cherrière et al., 2003), lysine availability (Batterham et al., 1986, Gdala et al., 1996) or a residual trace of alkaloids (Gdala et al., 1996). For the Amiga diet, the lower supply of digestible protein in the small intestine largely explained the drop in growth performance. The difference between the Lublanc and Boltensia diets is probably connected to the use of nutrients arriving in the blood stream. Parts of ingested and digested N retained by the pigs confirmed the better performance obtained with the Lublanc diet.
32The digestion of a-galactosides. The Boltensia diet supplied lower amounts of a-galactosides than white lupin-based diets (Table 11). In agreement with the findings reported by Cherrière et al. (2003), blue lupin was poorer in stachiose than the white varieties, but similar in its raffinose and verbascose content. Surprisingly, the caecal flows of a-galactosides were higher with the Boltensia diet, reflecting less degradation of these molecules due to less activity of the enzyme or lower fermentation of a-galactosides at the end of the small intestine. The major difference between the white lupin-based diets and the Boltensia diet stemmed from the physico-chemical properties of the fibres. It is possible that the transit time of the digesta was reduced with blue lupins because of a lower OM digestibility in the small intestine due to its high soluble fibre content. The resulting flow of the digesta was likely to limit the access of micro-organisms and to reduce the action time of the a-galactosidase. The difference between our results and those reported in the literature, suggesting that blue lupins are a better source of protein for growing pigs than white lupins, could be because these seeds had not been compared in the presence of an a-galactosidase supplement. Consequently, the lower amounts of a-galactosides contained in blue lupins was enough to make it preferable for use in pig feed, but once these molecules were neutralized, as in our study, white lupins resulted in a similar or higher zootechnical performance, depending on the cultivar.
4. Conclusions
33Our results suggest that a-galactosides are the main anti-nutritional factors in lupin seeds for growing and finishing pigs. Once these molecules were neutralized, it seemed that replacing SBM almost completely by lupin seeds not only did not reduce N retention in the pigs, but also improved carcass quality, particularly the fatty acid profile.
34In the presence of an a-galactosidase supplement, white lupin varieties (Lupinus albus cv. Amiga and Lublanc) led to a better zootechnical performance than blue lupin (Lupinus angustifolius var. Boltensia). Although the reasons were not clear, it appeared that, despite better intestinal digestibility, proteins supplied by the Boltensia diet was less used compared to proteins supplied by white lupins. The fact that blue lupins are recommended more often for pig feed than white lupins probably stems from the low a-galactosides content and from the fact that lupin species were not compared in the presence of a-galactosidase. It would be interesting to confirm these results using a larger number of lupin species and to investigate methods other than the one involving an enzymatic supplement, because the cost of including this supplement is still too high to be competitive in practical conditions.
35Overall, economical considerations are often difficult because the interests of using lupin seed in feeding do not only concern the animal aspects (performances, products quality improvements) but also the crop aspects, due to the fact that lupin is a good main crop, not requiring N fertilizers and having a low energy cost. These last aspects are not easily calculable. In a previous paper, we estimated the price of lupin seed lower to that of soybean meal only when lupin is grown by the breeders (Froidmont et al., 2004). In the future, lupin will probably be essentially used in sectors producing high quality products (good fatty acid profile, low w6/w3 ratio and high technological specifications) or favouring the utilization of farm products free from GMO or contaminants.
36Acknowledgement
37This research was subsidized by the Ministère de la Région Wallonne (MRW-DGA-IG3), Direction de la Recherche, Namur, Belgique. The authors wish to thank L. Nieuwenhuis, Novozymes SA, Bagsvaerd (Denmark) for the gift of the a-galactosidase and P. Bosh, A. Colinet, B. Grégoire, G. Jean, C. Malburny, A. Pire and JC. Pichon for their technical assistance.
Bibliographie
AFNOR NF V 04-401 (2001). Viande, produits à base de viande et produits de la pêche - Détermination de l’humidité. Paris : Association française de Normalisation.
AFNOR NF V 04-402 (2001). Viandes, produits à base de viande et produits de la pêche – Détermination de la teneur en matières grasses totales. Paris : Association française de Normalisation.
AFNOR NF V 04-403 (2001). Viandes, produits à base de viande et produits de la pêche – Détermination de la teneur en matières grasses libres. Paris : Association française de Normalisation.
AFNOR V 18-120 (1997). Aliments des animaux. Dosage de l’azote, méthode par combustion (DUMAS). Paris : Association française de Normalisation.
AOAC (1990). Official methods of analysis. Arlington, VA: Association of Official Analytical Chemists.
Bach Knudsen K., Gonzalez E. (2004). Chemical and nutritional characterisation of white (Lupinus albus L.) and blue (Lupinus angustifolius L.) lupins. 5th European Conference on grain Legumes and 2nd International Conference on Legume Genomics and Genetics, Dijon, France.
Batterham ES., Andersen LM., Burnham BV., Taylor GA. (1986). Effect of heat on the nutritional value of lupin (Lupinus angustifolius), seed meal for growing pigs. Br. J. Nutr. 55, p. 169–177.
Carré B., Brillouet JM., Thibault JT. (1985). Characterisation of polysaccharides from white lupin (Lupinus albus L.) cotyledons. J. Agric. Food Chem. 33, p. 285–292.
Cherrière K., Albar J., Noblet J., Skiba F., Granier R., Peyronnet C. (2003). Utilisation du lupin bleu (Lupinus angustifolius) et du lupin blanc (Lupinus albus) par les porcelets en post-sevrage. Journées Recherche Porcine 35, p. 97–104.
Collomb M., Buhler T. (2000). Analyse de la composition en acides gras de la graisse de lait. I. Optimisation et validation d’une méthode générale à haute résolution. Mitt. Lebensm. Hyg. 91, p. 306–332.
CVB. (1999). Veevoedertafel. Centraal Veevoederbureau. Runderweg 6. Postbus 2176. 8203 AD Lelystad. Pays-Bas.
Delzenne N., Robertfroid MB. (1994). Physiological effects of nondigestible oligosaccharides. Leb. Wis. Tech. 27, p. 1–7.
Dunshea FR., Gannon NJ., van Barneveld RJ., Mullan BP., Campbell RG., King RH. (2001). Dietary lupins (Lupinus angustifolius and Lupinus albus) can increase digesta retention in the gastrointestinal tract of pigs. Aust. J. Agric. Res. 52, p. 593–602.
Folch J., Lees M., Sloane-Stanley GH. (1957). A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226, p. 497–509.
François E., Thill N., Théwis A. (1978). Méthode rapide de dosage de l’oxyde de chrome dans les aliments, les fèces et les contenus digestifs par titrage après oxydation nitroperchlorique. Ann. Zootech. 27, p. 355–361.
Froidmont E., Bartiaux-Thill N. (2004). Suitability of lupin and pea seeds as a substitute for soybean meal in high-producing dairy cow feed. Anim. Res. 53, p. 475–487.
Gdala J., Jansman AJM., van Leeuwen P., Huisman J., Verstegen MWA. (1996). Lupins (L. luteus, L. albus, L. angustifolius) as a protein source for young pigs. Anim. Feed Sci. Technol. 62, p. 239–249.
Gdala J., Jansman AJM., Buraczewska L., Huisman J., van Leeuwen P. (1997). The influence of a-galactosidase supplementation on the ileal digestibility of lupin seed carbohydrates and dietary protein in young pigs. Anim. Feed Sci. Techn. 67, p. 115–125.
Gdala J., Smulikowska S., Zdunczyk Z., Pastuszewska B. (1999). Nutrients and antinutrients in three lupin species grown in Poland. In van Santen E., Wink M., Weissmann S., Römer P. (Eds). 9th International Lupin conference, Klink/Müritz, Germany.
Girard JP., Bout J., Salort D. (1988). Lipides et qualités du tissu adipeux. Facteurs de variation. Journées Recherche Porcine 20, p. 255–278 (Paris. France).
ISO/IDF 182 (2002). International Standard - Preparation of fatty acid methyl esters.
Lebret B., Mourot J. (1998). Caractéristiques et qualité des tissus adipeux chez le porc. Facteurs de variation non génétiques. INRA Prod. Anim. 11 (2), p. 131–143.
Minitab Version 13. (2000). Minitab User’s Guide.
Mourot J. (2001). Mise en place des tissus sous-cutané et intramusculaire et facteurs de variation quantitatifs et qualitatifs chez le porc. INRA Prod. Anim. 14 (5), p. 355–363.
Noblet J., Le Goff G. (2001). Effect of dietary fibre on the energy value of feeds for pigs. Anim. Feed Sci. Techn. 90, p. 35–52.
Rahman MH., Hossain I., Moslehuddin (1997). Nutritional evaluation of sweet lupin (Lupinus angustifolius): net protein utilization (NPU), nitrogen balance and fractionation studies. Br. J. Nutr. 77, p. 443–457.
Roth-Maier DA., Böhmer BM., Roth FX. (2004). Effects of feeding canola meal and sweet lupin (L. luteus, L. angustifolius) in amino acid balanced diets on growth performance and carcass characteristics of growing-finishing pigs. Anim. Res. 53, p. 21–34.
Salgado P., Freire JPB., Mourato M., Cabral F., Toullec R., Lallès JP. (2002). Comparative effects of different legume protein sources in weaned piglets: nutrient digestibility, intestinal morphology and digestive enzymes. Livest. Prod. Sci. 74, p. 191–202.
UNIP (2002). Statistiques 2002. Paris, France : Union nationale interprofessionnelle des plantes riches en protéines.
Van Soest PJ., Robertson JB., Lewis A. (1991). Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, p. 3583–3597.
Veldman A., Veen WAG., Barug D., van Paridon PA. (1993). Effect of a-galactosides and a-galactosidase in feed on ileal piglet digestive physiology. J. Anim. Physiol. Anim. Nutr. 69, p. 57–65.
Warnants N., Van Oeckel MJ., De Paepe M. (2001). Fat in pork: image. Dietary modification and pork quality. Pig News Infor. 22 (4), p. 107N-113N.
Wiggins HS. (1984). Nutritional value of sugars and related compounds undigested in the small gut. Proc. Nutr. Soc. 43, p. 69–75.