
Flora and fauna from a new Famennian (Upper Devonian) locality at Becco, eastern Belgium
Abstract
The Becco locality (Liège province), belongs to the Theux tectonic window and represents a proximal, probably fluvial, environment corresponding to a channel infill. We present here a preliminary report of the fossil assemblage discovered at the locality. The Becco site has yielded a diverse flora of early seed plants including Moresnetia zalesskyi, Dorinnotheca streeli and Condrusia sp. This assemblage, characteristic of the Belgian Famennian, highlights the diversity of early spermatophytes in the country. Becco has also delivered a rich vertebrate fauna with antiarch, groenlandaspid and phyllolepid placoderms, diplacanthiform acanthodians, as well as actinopterygians and various sarcopterygians. The fossiliferous assemblage of Becco resembles those of several Devonian tetrapod-bearing localities, including that of Strud in Belgium, and could therefore provide a favorable palaeoecological setting in the search for early tetrapods.
1. Introduction and geological context
1In Belgium, Famennian rocks crop out extensively across the Dinant Synclinorium (allochtonous), Haine-Sambre-Meuse Overturned Thrust Sheets (former southern limb of the Namur Synclinorium; see Belanger et al., 2012, for discussion about this Variscan structural element), Vesdre area (lateral equivalent to the latter), Theux Tectonic Window (autochtonous) and Brabant Parautochton (former northern limb of the Namur Synclinorium, parautochtonous). The Famennian shows a regressive trend from open marine shales of the Famenne Group (lower Famennian) to fluvio-lagunal siltstones, sandstones and dolomites of the Upper Famennian Evieux Formation (Thorez & Dreesen, 1986). The litho- and biostratigraphy of the stage are well known, thanks to the seminal works of Thorez and colleagues (Thorez et al., 1977; Thorez & Dreesen, 1986, 2002; Thorez et al., 2006), which were mainly dedicated to the Condroz area (eastern Dinant Synclinorium). The Famennian lithostratigraphy of the Haine-Sambre-Meuse Overturned Thrust Sheet (sensu Belanger et al., 2012), Vesdre area and Theux Tectonic Window, even if similar, remains relatively poorly known. In comparison to contemporaneous deposits exposed in the Dinant Synclinorium, the Upper Famennian deposits known from these Variscan structural units are more proximal and the fluvial and lagunal facies are commonly more developed (Thorez et al., 1988).
2This paper describes a recently discovered Upper Famennian locality in the Theux Tectonic Window, which has yielded a particularly rich and diverse flora and vertebrate fauna. The Becco outcrop was discovered by two of the authors (J.-M. M. and B. M.) during the geological survey undertaken as part of the geological modernization program of the Geological map of Wallonia (Louveigné-Spa sheet). The outcrop is situated on a road embankment close to Becco hamlet in the vicinity of Theux (Liège province) (Fig. 1). The stratigraphic succession exposed at Becco is reduced and consists mainly of green shaly siltstone belonging to the Crupet Member (Evieux Formation). The Becco site is a limited road-cut exposure and fossil excavation is currently only possible by picking blocks laterally. Most of the fish fossil layers are prone to strong fracturing making fossil sampling difficult. Despite limited fieldwork so far, the Becco outcrop has already yielded a fossil flora characterized by early seed plants and a very diverse fauna including placoderms, acanthodians, actinopterygians, and sarcopterygians.
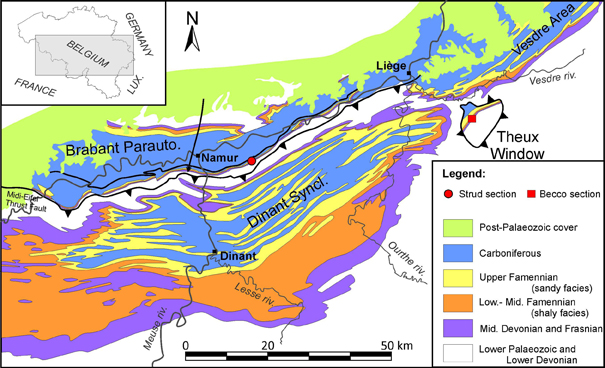
3Figure 1. Geological map of Southern Belgium with the location of the Becco (red square) and Strud (red circle) localities (redrawn after de Béthune, 1954).
2. Material and methods
4All specimens were mechanically prepared. Plants and some fish remains were photographed under water immersion to enhance certain morphological characters.
2.1. Institutional abbreviations
5IRSNB, Institut Royal des Sciences Naturelles de Belgique, Brussels; ULg, Université de Liège.
2.2. Anatomical abbreviations
6a.b, annular bourrelet/thickening; ADL, anterior dorsolateral plate; AVL, anterior ventrolateral plate; dlr, dorso-lateral ridge; emb.ADL, embayment of MD for ADL; emb.PDL, embayment of MD for PDL; f.ax, axillary foramen; f.art, articular fossa of ADL; lc, main lateral sensory line groove; MD, median dorsal plate; MV, median ventral plate; oa.ADL, overlap area for ADL; oa.MD, overlap area for MD; oa.MV, overlap area for MV; oa.PDL, overlap area for PDL; oa.PMD, overlap area for PMD; PDL, posterior dorsolateral plate; PMD, posterior median dorsal plate; PVL, posterior ventrolateral plate; pro, processus obstans; tsb, triangle-shaped bump; vlr, ventro-lateral ridge.
3. Systematic palaeontology
3.1. Early seed plants
7Plant fossils are rare in this locality, mostly consisting of isolated short lengths of isotomously or anisotomously branched axes. Two types of fertile specimens have been discovered: (?micro) sporangiate structures that are currently under study at Liège University and early seeds that are described here. Several types of early seed plants are present; all belong to the hydrasperman type of early seeds (sensu Rothwell, 1986): the apical part of their megasporangium is modified into a structure interpreted as favouring the pollination process. The early seed plants collected at Becco are representative of 3 morphotypes (Moresnetia, Dorinnotheca and Condrusia types) of the 5 identified by Prestianni (2005) and illustrated by Prestianni & Gerrienne (2010, fig. 3).
3.1.1. The Moresnetia-type
8Fig. 2A-C
9The Moresnetia-type includes seven different genera, collected from at least 12 localities distributed on the palaeocontinent Laurussia (Prestianni & Gerrienne, 2010). Early seeds of the Moresnetia-type are enclosed in a cupule, consisting of a cup-like bunch of dichotomous branch tips. The Moresnetia-type is characterized by a four-unit cupule, formed by two successive cruciate dichotomous divisions (Prestianni, 2005; Prestianni & Gerrienne, 2010). The cupule defines a well circumscribed space where one to four seeds (exceptionally six) are found. The hydrasperman megasporangium is surrounded by several variably fused integumentary lobes.
10Moresnetia zalesskyi Stockmans (1948), emend. Fairon-Demaret & Scheckler (1987) is commonly collected from various localities in the Late Devonian Evieux Formation from Belgium, namely Assesse, Hun-Annevoie, Hamois, Moresnet (Stockmans, 1948), Dorinne, Evrehailles (Fairon-Demaret & Scheckler, 1987) and Strud (Stockmans, 1948; Prestianni et al., 2007). The plant had a shrubby habit, with a main stem supporting large, profusely branched, lateral systems that distally bear numerous cupules (Serbet & Rothwell, 1992; Prestianni et al., 2007). Those lateral systems are generally considered homologous to large leaves, but they do not show any laminated parts.
11Five specimens of Moresnetia zalesskyi have been collected (Fig. 2A-C) at Becco. They consist of the distal part of the seed-bearing system. Axes are 0.5-1 mm wide and show slightly anisotomous branching. Each axis is terminated by a cupule. Cupules are 8-10 mm long and 5-8 mm wide. One badly preserved seed is visible on figure 2C.

12Figure 2. Plant fossils from Becco. A. ULgBecco 6, Moresnetia zalesskyi Stockmans (1948), emend. Fairon-Demaret & Scheckler (1987). Scale bar equals 1 cm. B. ULgBecco 3-1, Moresnetia zalesskyi Stockmans (1948), emend. Fairon-Demaret & Scheckler (1987). Scale bar equals 1 cm. C. Moresnetia zalesskyi Stockmans (1948), emend. Fairon-Demaret & Scheckler (1987): enlargement of Fig. 2B, a badly preserved seed is visible at arrow. Scale bar equals 2 mm. D. ULgBecco 2-1, Dorinnotheca streelii Fairon-Demaret (1996a). Scale bar equals 2 mm. E. Dorinnotheca streelii Fairon-Demaret (1996a): enlargement of Fig. 2E showing the putative collarette cupule. Scale bar equals 1 mm. F. ULgBecco 3-2, Condrusia sp. Scale bar equals 2 mm. G. ULgBecco 2-2, Condrusia sp. Scale bar equals 2 mm.
3.1.2. The Dorinnotheca-type
13Fig. 2D-E
14The Dorinnotheca-type includes a single species, Dorinnotheca streelii (Fairon-Demaret, 1996a). According to the original description, the external structure of the seed is interpreted as a large, highly dissected cupule composed of 8 proximally fused parts, with at least 40 distal segments (Fairon-Demaret, 1996a). This morphology has been considered suggesting adaptations for wind dispersal (Prestianni & Gerrienne, 2010). A single seed is present in each cupule. Before this work, Dorinnotheca was only known from three Belgian localities (Angleur, Dorinne, Trooz; Fairon-Demaret, 1996a). The gross morphology of the plant is unknown.
15A very well-preserved specimen of Dorinnotheca streelii has been found at the Becco locality (Fig. 2D). It is borne at tip of a 0.5-1.0 mm wide, isotomously branched axis segment; the seed itself (external structure included) is 10 mm long and 8 mm wide. The specimen suggests an alternative interpretation for the external structure of the seed. It shows a small subterminal structure (Fig. 2E) that exhibits striking similarities for the diminutive cupule of Pseudosporogonites Stockmans, another early seed plant from Belgium, UK and USA (Prestianni et al., 2013). In this genus, the cupule, called collarette by Prestianni et al. (2013) is a very small organ, located at the very base of the seed. The specimen of Dorinnotheca from Becco shows a comparable structure (Fig. 2E) that might also be a collarette cupule. It is noticeable that a similar collarette is visible on the holotype specimen (Fairon-Demaret, 1996a, pl. I, fig. 1, at the end of the pedicel of seed c, lower part of the figure). If our interpretation of this organ is correct, it suggests that the highly dissected structure surrounding the Dorinnotheca seed is the tegument, not the cupule. This has however to be confirmed by the discovery of other comparable specimens.
3.1.3. The Condrusia-type
16Fig. 2F-G
17The Condrusia-type includes the genus Condrusia Stockmans (Stockmans, 1948) only. Three species are known (Stockmans, 1948; Prestianni & Gerrienne, 2006); they are currently being re-evaluated at Liège University. Abundant Condrusia specimens come from Belgian localities, e.g. Assesse, Dorinne, Hamois, Villers-le-Temple, Strud (Stockmans, 1948; Fairon-Demaret, 1996b; Prestianni et al., 2007). The cupule of the Condrusia-type is borne distally at the tip of a narrow, often bent, axis segment. The cupule is a flat structure consisting of two leaf-like segments closely adpressed against each other. The cupule outline is characteristically trapezoidal or diamond-shaped. Each cupule contains one bottle-shaped seed. The gross morphology of the plant is unknown.
18Several Condrusia specimens have been collected at Becco; two of them are illustrated here (Fig. 2F-G). They are approximately 6 mm long and wide. Both show the radially symmetrical seed and possible trace of the vascularisation of the cupule segments.
3.2. Fauna
19Preliminary investigations at the Becco site have yielded numerous placoderm remains but also isolated elements of acanthodian, actinopterygian and sarcopterygian fishes. Such a fish assemblage with rich osteichthyan remains is quite scarce in the Famennian of Belgium and the Becco site therefore deserves special attention due to the strong potential of this locality.
20Class Placodermi McCoy, 1848
21Order Euantiarcha Janvier & Pan, 1982
22Suborder Asterolepidoidei Miles, 1968
23Family Remigolepidae Stensiö, 1931
24Genus Remigolepis Stensiö, 1931
25Type species.—Remigolepis incisa (Woodward, 1900). Ymer Island, south of Dusen Fjord, East Greenland, Famennian.
26Remigolepis cf. R. durnalensis Olive, 2015
27Fig. 3A-D
28A badly-preserved anterior dorsolateral plate is exposed in visceral view (Fig. 3A). It displays a strong processus obstans (pro) and an extended articular fossa (f.art). The overlap area for the posterior dorsolateral plate (oa.PDL) is partly visible.
29A posterior dorsolateral plate (Fig. 3B-C) is exquisitely preserved. Bone was removed in order to access both external and visceral views. It corresponds to the description of R. durnalensis made by Olive (2015), which could be slightly enlarged by some new features: 1) a narrow area overlapped by the posterior median dorsal plate (oa.PMD), 2) a clearly visible main lateral sensory line groove (lc) running alongside the convex ventral margin of the lateral lamina as suggested by Olive (2015) and 3) a short area overlapped by the anterior dorsolateral plate (oa.ADL).
30An almost complete ventral trunk shield is displayed in visceral view (Fig. 3D). Bone of both the anterior ventrolateral (AVL) and the left posterior ventrolateral (PVL) plates is preserved whereas only the imprint of the external surface of the right posterior ventrolateral plate is present. The lateral lamina of the right anterior ventrolateral plate is crushed. It is distinguishable from the ventral lamina by the ventrolateral ridge (vlr). The ventral lamina is elongate with a moderate anterior notch (compared to that of R. walkeri for example; Johanson, 1997), usually for the paired semilunars which are absent in this specimen. The axillary foramen (?f.ax) is difficult to observe. The crista transversalis interna anterior is slightly marked due to the compression. The median ventral plate (MV) is missing, allowing the observation of the long and narrow overlapping areas on the anterior ventrolateral plates (oa.MV). The space occupied by the median ventral plate is relatively small and therefore contradicts the assumption made by Olive (2015) for R. durnalensis, i.e. “the median ventral plate should occupy a large place in the ventral shield”.
31The ornament is made of vermiculate, coarse, short ridges and of large tubercles.
32Remark 1—The antiarch material found at Becco is assigned to Remigolepis because of the sigmoid suture of the posterior dorsolateral plate with the anterior median dorsal plate, i.e. the posterior dorsolateral plate is overlapped by the anterior median dorsal plate anteriorly, whereas the posterior dorsolateral plate overlaps the anterior median dorsal plate posteriorly.
33The material of Remigolepis from Becco is assigned to Remigolepis cf. R. durnalensis because of the processus obstans well developed and the extended articular fossa of the anterior dorso-lateral plate, as in the type material of R. durnalensis. The vermiculate and tuberculate ornament also resembles that of R. durnalensis type material. The Famennian age for both localities pleads also for the comparison. However, there is currently not enough material to affirm with certainty that R. durnalensis is found at Becco.
34Remark 2—The material found in Becco is larger than the type material described by Olive (2015) from Durnal and certainly corresponds to more mature specimens. The low lateral lamina of the posterior dorsolateral plate seems to confirm what Olive (2015) suggested concerning the compression of the trunk armour with the growth of the animal.

35Figure 3. Placoderm fish content from Becco. A. IRSNB P.9317a, Remigolepis cf. R. durnalensis, ADL in internal view. B. IRSNB P.9318a, Remigolepis cf. R. durnalensis, PDL in external view. C. IRSNB P.9318b, counterpart of IRSNB P.9318a, Remigolepis cf. R. durnalensis, PDL in internal view. D. IRSNB P.9319, Remigolepis cf. R. durnalensis, ventral trunkshield in internal and external views. E. IRSNB P.9320a, ?Phyllolepis sp., AVL in internal and external views. F. IRSNB P.9321, Groenlandaspis potyi, MD and ADL in internal and external views. G. IRSNB P.9322a, Groenlandaspis potyi, PDL in internal and external views. H. IRSNB P.9323a, ?Groenlandaspis sp., putative MD in external view.
36Order Arthrodira Woodward, 1891
37Suborder Actinolepidoidei Miles, 1973
38Infraorder Phyllolepidida Stensiö, 1934
39Family Phyllolepididae Woodward, 1891
40Genus Phyllolepis Agassiz, 1844
41Type species.—Phyllolepis concentrica Agassiz, 1844. Perthshire and Rosebrae beds, Morayshire, Scotland, Famennian.
42?Phyllolepis sp.
43Fig. 3E
44Only an anterior ventrolateral plate is observable in external view (Fig. 3E). The length/width index is of 1.4 (21 mm/15 mm). The lateral margin is one third shorter than the medial margin. The ornament is made of well-defined parallel lines, less spaced posteriorly.
45Remark 1—The distinctively ridged dermal ornament is characteristic of Phyllolepidae. A sole anterior ventrolateral plate does not permit generic attribution. However we assigned cautiously the anterior ventrolateral plate of Becco to the genus Phyllolepis because of the abundance of this genus in other Famennian localities from Belgium, e.g. Strud, Chèvremont and Evieux (Lohest, 1888; Leriche, 1931). Moreover, to date, this is the only Phyllolepididae genus found in Euramerica.
46Suborder Phlyctaenioidei Miles, 1973
47Infraorder Phlyctaenii Miles, 1973
48Family Groenlandaspididae Obruchev, 1964
49Genus Groenlandaspis Heintz, 1932
50Type species.—Groenlandaspis mirabilis Heintz, 1932. East Greenland, Upper Famennian.
51Groenlandaspis potyi Olive et al., 2015
52Fig. 3F-G
53The material consists of a median dorsal plate in connection with an anterior dorsolateral plate (Fig. 3F) and of an isolated posterior dorsolateral plate (Fig. 3G). The shape of the median dorsal plate resembles a rough equilateral triangle, as in the type material (Olive et al., 2015), but the gentle posteroventral slope observed in the type material is not preserved in this specimen. The tip of the median dorsal plate, missing in the type material, appears to be blunt in the present specimen. The asymptotic lines of the anterior and posterior edges form an angle of 75°. The ventral margin of the plate is deeply indented by the posterior dorsolateral plate (emb.PDL).
54The anterior dorsolateral plate (in visceral view; Fig. 3F) is badly preserved with the posterior part and articular condyle missing. The limits with the median dorsal and possibly the posterior dorsolateral plates are blurry.
55The posterior dorsolateral plate is preserved in visceral view for the ventral part and in external view for the dorsal part (Fig. 3G). The high flexure of the main lateral sensory line groove (lc) is discernible; it forms an angle of 80°. Posteriorly, the groove leaves the PDL at the first ventral third of the plate height. Those features were not observable on the holotype of G. potyi and enlarge the description of the species. The external “triangle-shaped bump” (tsb) (Olive et al., 2015), above the lateral sensory line groove flexure and extending to the posterior margin, is observable as for the type material, and the annular thickening is clearly visible (a.b). The overlap area for the anterior dorsolateral plate (oa.ADL) is extended. Because the posterodorsal angle is not entirely preserved, it is impossible to say if the overlap area for the median dorsal plate (oa.MD) reaches the posterodorsal apron (a diagnostic character of Groenlandaspis potyi; Olive et al., 2015).
56No ornament is observable in the limited amount of material showing the external sides. The large tubercles, present on the anterodorsal corner of the posterior dorsolateral plate (Fig. 3G), belong very certainly to a plate of Remigolepis cf R. durnalensis present below the posterior dorsolateral plate.
57Remark 1—This material is assigned to the taxon Groenlandaspis potyi because of the equilateral triangle-shaped median dorsal plate and the triangle-shaped bump on the external surface of the posterior dorsolateral plate.
58Remark 2—This Groenlandaspis potyi material from Becco is smaller than the type material described by Olive et al. (2015) from Hun-Annevoie and Durnal and certainly corresponds to juvenile or less mature specimens.
59?Groenlandaspis sp.
60Fig. 3H
61A triangular element, possibly attributable to a median dorsal plate of Groenlandaspis sp., was also found in Becco. The ventral part is not preserved but the plate seems lower than that of G. potyi. Its apex is located slightly posteriorly from the midlength (while it is at midlength in G. potyi). The embayments for the posterior dorsolateral (emb.PDL) and anterior dorsolateral (emb.ADL) plates are not as angular as in G. potyi (though the distance between the tip of the median dorsal plate and the tip of the embayments is identical between this specimen and G. potyi). The embayment for the posterior dorsolateral plate is slightly higher than the embayment for the anterior dorsolateral plate. The tip of the plate is pointed. The anterior and posterior edges form an angle of 81°. The anterior edge is covered with pointed tubercles. There is no observable ornamentation.
62Class Acanthodii Owen, 1846
63Order Diplacanthiformes Berg, 1940
64Family Diplacanthidae Woodward, 1891
65Diplacanthidae indet.
66Fig. 4A
67IRSNB P.9325 is a massive, longitudinally ridged spine, measuring 16 mm x 2 mm. Its proportions are different (L/w = 8.5) from IRSNB P.9327 and 9324. It displays up to 16 ridges on the lateral side, a polygonal section with a prominent ridge on the trailing edge and the inserted part (right on Fig. 4A3) displays well marked ridges, considered a diplacanthiform character. On other diplacanthiform spines, grooves and ridges seem more marked on the distal part of the spine, e.g. in Milesacanthus (see Young & Burrow, 2004, fig. 5) or in “Onchus” concinnus and “O.” marginalis (Gross, 1933, p. 19, pl. I: 3-6). On the contrary here, the distal part of the spine, visible on the counterpart (Fig. 4A2), is laterally smooth but surely worn. The limit between the outside (left on Fig. 4A1) and the more reduced inside parts (right on Fig. 4A1; 4A3) is oblique.
68Remark 1—Diplacanthidae possessed spines ornamented with smooth longitudinal ridges paralleling the leading edge of the spine (Young & Burrow, 2004, p. 25), like IRSNB P.9325.
69Remark 2—In this study, this spine was first attributed to Devononchus cf D. marginalis but the specimen is more ridged and the histology is unknown. It is very similar to spines from Strud.

70Figure 4. Acanthodian fish content from Becco. A. IRSNB P.9325a and b, Diplacanthidae indet., (A1) IRSNB P.9325a, lateral view, (A2) close-up of the distal part of the counterpart (IRSNB P.9325b), (A3) close-up of the limit between free part and inserted part. B. IRSNB P.9326a, Diplacanthidae? indet., (B1) lateral view, (B2) close-up of the central part. C. IRSNB P.9324, Acanthodii indet., lateral view. D. IRSNB P.9327, Acanthodii indet. posterior view?. E. IRSNB P.9328a, Acanthodii indet. (E1) trailing edge view, (E2) close-up of the distal part.
71Diplacanthidae? indet.
72Fig. 4B
73Spine IRSNB P.9326 is flattened (possibly by compression, 4B1) and is about 2 cm long. It shows a width of 1.18 mm. It displays more than 10 very tiny longitudinal ridges (Fig. 4B2) underlined by black coloration. They are laterally visible in the counterpart and also on the leading edge.
74Remark 1—IRSNB P.9326 is attributed with caution to the family diplacanthidae since the ridges are not well marked.
75Acanthodii indet.
76Fig. 4C
77IRSNB P.9324 is a slender spine measuring 9 mm x 0.4 mm set in a micaceous green sandstone. It displays the same proportions as spine IRSNB P.9327 (L/w = 20). The section is triangular and the surface is smooth. It could be “Onchus” tenuispinus, first described in Ketleri (Latvia) and characterized by the absence of longitudinal ridges (Gross, 1933, p. 21). Later, Gross (1940) transferred some Onchus species (like O. concinnus, O. laevis and O. tenuispinus) to the genus Devononchus thanks to histological data, but the scarcity of material did not allow to make thin sections. Devononchus tenuispinus is one of the few taxa with smooth rounded spines (Burrow, 2007).
78Fig. 4D-E
79Spine IRSNB P.9327 (Fig. 4D) is slender and measures 23 mm x 1.1 mm (L/l = 20.91). It is broken across the width and for most of the spine no external surface is visible. However, some ridges at the base are observable. The spine displays a triangular section.
80Spine IRSNB P.9328 (Fig. 4E) has a length of 14 mm and is exposed in posterior view (Fig. 4E1). Some areas have been shifted by compression. The spine is embedded in a green micaceous sandstone like IRSNB P.9324. Longitudinal ridges are visible in the depressed part (Fig. 4E2) and in the counterpart. Proportions are like those of spine IRSNB P.9326 (L/w = 14).
81Megaclass Osteichthyes Huxley, 1880
82Fig. 5A
83A slender isolated gular plate presents an ornamentation of numerous and fine longitudinal ridges. The transverse pit-line is clearly visible in the middle of the plate. Gular plates are unfortunately not diagnostic within the osteichthyans and it could belong either to the actinopterygian or the actinistian clades.

84Figure 5. Osteichthyan fish content from Becco. A. IRSNB P.9329a, Osteichthyes gen. et sp. indet., gular in external view, photographed under water immersion. B. IRSNB P.9330, Holoptichiid porolepiform, scale in external view. C. IRSNB P.9331a, cf. Soederberghia sp., scale in internal view. D. IRSNB P.9332, cf. Soederberghia sp., scale in internal view. E. IRSNB P.9332, cf. Soederberghia sp., same as D with bone removed in order to show the ornamentation of the external side of the scale. F. IRSNB P.9333, cf. Tristichopteridae, isolated tooth. G. IRSNB P.9334a, cf. Tristichopteridae, slab with numerous scattered scales, most probably from the same individual. H. close-up on IRSNB P.9334a, cf. Tristichopteridae, a small-sized scale. I. IRSNB P.9335, cf. Tristichopteridae, scale in internal view. J. IRSNB P.9336a, Tetrapodomorpha indet., unidentified dermal element.
85Superclass Sarcopterygii Romer, 1955
86Subclass Dipnomorpha Ahlberg, 1991
87Order Porolepiformes Jarvik, 1942
88Family Holoptychiidae Owen, 1860
89Fig. 5B
90Isolated scales of a holoptychiid porolepiform are abundant in Becco. They are easily recognizable by their very large size, the non-cosmine covered free region ornamented with large anastomosed ridges, and the absence of a drop-shaped boss on their inner side (Jarvik, 1972). Holoptychiid scales are the most common sarcopterygian remains in the Famennian localities of Belgium, which is also the case for almost all sarcopterygian-bearing localities of the world known as proximal to true continental environments (estuarine, fluviatile, lacustrine; e.g., Lebedev et al., 2010).
91Order Dipnoiformes
92Family Rhynchodipteridae Moy-Thomas, 1939
93cf. Soederberghia Lehman, 1959
94Type species.—Soederberghia groenlandica Heintz, 1932. East Greenland, Upper Famennian.
95Fig. 5C-E
96The Becco site has yielded large, round and thin scales, similar in size and shape to holoptychiid scales but with a very different ornamentation. The inner side shows a well-defined punctuated area and well-marked concentric growth lines, as well as an absence of a central drop-shaped knob. These features are clearly present on the inner side of the scales of the large lungfish Soederberghia groenlandica from East Greenland (Lehman, 1959, pl. 20A; Friedman, 2007, fig. 8). Strikingly it seems that the punctuated area on the inner side delimits the opposite free area of the external side. The free area is ornamented with numerous very thin and parallel anastomosed ridges. This distinctive ornamentation is known in the lungfishes Soederberghia groenlandica (Lehman, 1959; Friedman, 2007), Soederberghia simpsoni (Ahlberg et al., 2001), Rhynchodipterus elginensis (Säve-Söderbergh, 1937) Fleurantia denticulata and Scaumenacia curta (Cloutier, 1996).
97Large lungfishes from Belgium were previously only recorded in the Famennian localities of Strud, Namur province, and Modave, Liège province, with the genus Soederberghia (Clément & Boisvert, 2006).
98Subclass Tetrapodomorpha Ahlberg, 1991
99Order Osteolepiformes Berg, 1937
100cf. Tristichopteridae Cope, 1889
101Fig. 5F-I
102Isolated teeth and scales of large tetrapodomorph fishes were also found at Becco. Teeth are quite long and large, laterally flattened, and with cutting edges (Fig. 5F). A few deep but short striae are present from the base to mid-length of the tooth. Scales are abundant and sometimes found scattered on the same slab (Fig. 5G). The small-sized scales (less than 1 cm in diameter) are more or less hexagonal (Fig. 5H) whereas the large ones (around 1 cm) are pear-shaped (Fig. 5I). A distinct drop-shaped bump is present on their internal side, central but generally positioned closer to the anterior edge. The posterior margin is slightly indented. Growth ridges parallel to the margin are usually present on the inner side. When the basal layer of the scale is missing (broken or not mineralized) an intense network of radial circuli is visible (Fig. 5H).Isolated rhizodontid and tristichopterid scales are hardly distinguishable from each other. However the presence of quite large pear-shaped scales with fringed posterior margins strongly supports a tristichopterid determination for this scattered material from Becco.
103Two large tristichopterids are known in the Famennian of Belgium, the genera Eusthenodon and Langlieria (Clément et al., 2009). They can sometimes be found associated (Clément, 2002). It is unfortunately not possible to discriminate these two taxa on the basis of isolated teeth or scales.
104Tetrapodomorpha indet.
105Fig. 5J
106An isolated dermal element presents an ornamentation of pronounced anastomosed ridges characteristic for tetrapodomorphs. The bone is complete and quite thick in relation to its small size. The overall shape is rectangular and anastomosing ridges cover all external surface. The internal side presents long and thin parallel striae. These striae and the absence of overlap area imply that this dermal bone is probably not an isolated scale of a non cosmine covered tetrapodomorph although it recalls the overall scale morphology of Litoptychius (Schultze & Chorn, 1998; Coates & Friedman, 2010) or Glyptopomus (Jarvik, 1950). This small bone could rather be an element of the skull, such as an isolated nasal or supraorbital, of a large tetrapodomorph.
4. Discussion
4.1. Taphonomy of the Becco site
107According to local and regional stratigraphical and sedimentological observations, the facies indicates a fluvial environment and the fossiliferous horizon most likely corresponds to a channel infill.
108The plant fossils at the locality are generally well preserved, but of short length. This, together with their scarcity and their scattered distribution in the sediment, suggests a moderately long transport in a low-energy system and a rapid burial.
109The vertebrate remains were likely brought by currents and deposited as a lag deposit and now form a bone bed a few centimetre-thick. Except for the acanthodian remains, which are scarce and damaged, the fish material from Becco is well preserved, although composed of isolated dermal bones, scales and teeth. Some complete specimens could however be expected since placoderms have been found in partial anatomical connection (Fig. 3D), and since a slab with numerous osteolepiform scales (Fig. 5G), most probably from the same specimen, has been collected. The Famennian deposits of Belgium rarely provide vertebrate specimens in connection. Except the unique specimen of the actinopterygian Osorioichthys, described from the Belgian Lower Famennian (Taverne, 1997), displaying an anterior part very well preserved and some placoderms with dermal plates in connection, e.g. the head shield of Grossilepis rikiki from Moresnet (Olive, 2015) and the trunk shield of Groenlandaspis potyi from Hun-Annevoie (Olive et al., 2015), most vertebrate plates and bones are found isolated. It differs from some Carboniferous localities such as the Viséan marine conservation-Lagerstätte of Denée, where exquisitely preserved whole specimens of the actinopterygian Benedenius (Mottequin et al., 2015) are present.
4.2. The fossil plant assemblage in a global context
110The Becco fossil plant record is particularly scarce. All identifiable plant remains are spermatophytes. Three distinct morphotypes are present: Moresnetia, Dorinnotheca and Condrusia types. They highlight the diversity of this group in Belgium. The Famennian stage indeed records one of their earliest diversification events of the spermatophyte clade (Hilton, 1998; Prestianni & Gerrienne, 2010). Most, if not all, of the 17 Famennian seed plant species are reported from the equatorial belt (present day USA, Belgium, Great-Britain, Germany, Ireland and Russia). In this context, eastern Laurussia (present day Great-Britain, Ireland and Belgium), where 11 species have been described, seems to represent a region of high diversity. This situation has been recently discussed by Prestianni & Gerrienne (2015). They suggest that the high diversity level of early seed plants collected from eastern Laurussia might be explained by the particularly dry climatic conditions that were prevailing there. The “extreme” heterospory, characteristic of the seed habit, very likely gave the early spermatophytes an ecological advantage in dryer environments.
4.3. The fossil fish assemblage in the Belgian framework
4.3.1. Placoderms
111The placoderm assemblage from Becco (and at a larger extant the whole vertebrate assemblage) is clearly dominated by the antiarch Remigolepis cf. R. durnalensis. That species is also very common in the type locality of Durnal (Olive, 2015). Phyllolepis is a common genus in the Famennian deposits of Evieux, Chèvremont and Strud (Lohest, 1888; Leriche, 1931). In the latter, that genus is abundant whereas only one plate has been found in Becco. Concerning the groenlandaspididae material, only Hun-Annevoie and Durnal have delivered Groenlandaspis potyi (Olive et al., 2015). Becco is the only Belgian locality displaying more than one species of Groenlandaspididae.
4.3.2. Acanthodians
112Some acanthodian spines from Becco are considered as diplacanthidae spines since they are characterized by smooth longitudinal ridges paralleling the leading edge of the spine (Young & Burrow, 2004, p. 25). The material is worn and furthermore no thin sections were made due to the scarcity of spines in Becco locality, so diagnostic characters were difficult to emphasize.
113This material, only composed of isolated ridged and smooth spines, is comparable with that of the nearby Upper Famennian localities of Durnal and Strud; but Durnal includes also the mesacanthid Teneracanthus sp. (Derycke & Clément, 2013). No shark remain has been found in these three localities, i.e. Becco, Durnal and Strud, on the contrary to Modave, Rance and Dinant where chondrichthyan scales have been previously described from Famennian levels (Derycke & Chancogne-Weber, 1995; Derycke-Khatir, 2005). This absence may be explained by the very proximal location of Becco compare to the distal position of Modave, Rance and Dinant.
4.3.3. Osteichthyans
114The Becco osteichthyan assemblage is characteristic of the Famennian fauna from Belgium with numerous remains of holoptychiid porolepiform and tetrapodomorph fishes. A similar faunal composition is known in different Famennian vertebrate-bearing localities of the Namur and Liège provinces, e.g. Strud, Modave, Durnal, Spontin, Chèvremont (Lohest, 1888; Leriche, 1931; Clément, 2002; Clément & Boisvert, 2006; Clément et al., 2009).
4.4. A potential Famennian tetrapod-bearing locality
115If we ignore the Becco site, the only Famennian locality from Belgium having so far yielded antiarch, groenlandaspididae and phyllolepid placoderms is the tetrapod-bearing locality of Strud (Clément et al., 2004; Olive, 2015; Olive, pers. obs.). The association of Phyllolepis species with early tetrapods in the northern hemisphere is recurrent. Consequently, the occurrence of that placoderm may serve as an indicator for tetrapod-bearing localities (Young, 2005). This hypothesis is supported by the Strud locality (Namur province) and suggests that other Belgian localities, i.e. Chèvremont, Evieux and Becco, could potentially deliver tetrapod remains. The abundance of sarcopterygian remains in the Becco site and the similarity of its faunal composition with that of the tetrapod-bearing locality of Strud, i.e. acanthodians, placoderms and sarcopterygians, allow the possibility of future discovery of tetrapod remains in Liège province. Except for the acanthodian fauna, the vertebrate assemblage of the Becco site also resembles those of worldwide Devonian tetrapod-bearing localities, e.g. Red Hill in Pennsylvania (e.g. Daeschler et al., 2003), East Greenland (e.g. Blom et al., 2007) and is another argument pleading for the possible presence of tetrapods in the Becco site. Due to the scarcity of early tetrapod remains worldwide, a new tetrapod occurrence in the Famennian of Belgium would be an important step in our knowledge of the origin and terrestrialization of vertebrates during Late Devonian times.
5. Conclusions
116Preliminary investigations led in the Upper Famennian locality of Becco (Liège province, eastern Belgium) have shown that the fossil assemblage of this site is much diversified. Its flora composition includes early seed plants such as Moresnetia zalesskyi, Dorinnotheca streeli and Condrusia sp. whereas its vertebrate composition includes diplacanthiform acanthodians, antiarch, groenlandaspid and phyllolepid placoderms, as well as various porolepiform, dipnoan and tetrapodomorph sarcopterygians. This assemblage, together with the proximal environment of the locality, are similarities shared with contemporaneous tetrapod-bearing localities. Therefore, the Becco site is promising in the search for early tetrapods and in our understanding of the fish-tetrapod transition.
6. Acknowledgements
117We gratefully acknowledge the Belgian Federal Science Policy Office for the research financial support (Doctoral Fellow to S. O.). Ph. G. is a F.R.S.-FNRS Senior Research Associate. The authors thank the editor and both reviewers J. Long and C. Berry for their fruitful remarks. The photographs were made by W. Miseur (IRSNB, Belgium) for placoderms and osteichthyans. The help of S. Ventalon (University of Lille 1) for the acanthodian photographs was really appreciated. This paper is a contribution to the ANR TERRES 2010-BLAN-607-03 project.
7. References
118Agassiz, L., 1844-1845. Monographie des poissons fossiles du Vieux Grès Rouge ou système Dévonien (Old Red Sandstone) des Iles britanniques et de Russie, XXXVI. Imprimerie Petitpierre, Neuchâtel et Soleure, 171 pp.
119Ahlberg, P. E., 1991. A re-examination of sarcopterygian interrelationships, with special reference to the porolepiformes. Zoological Journal of the Linnean Society, 103(3), 241–287.
120Ahlberg, P. E., Johanson, Z. & Daeschler, E. B., 2001. The Late Devonian lungfish Soederberghia (Sarcopterygii, Dipnoi) from Australia and North America, and its biogeographical implications. Journal of Vertebrate Paleontology, 21(1), 1–12.
121Belanger, I., Delaby, S., Delcambre, B., Ghysel, P., Hennebert, M., Laloux, M., Marion, J.-M., Mottequin, B. & Pingot, J.-L., 2012. Redéfinition des unités structurales du front varisque utilisées dans le cadre de la nouvelle Carte géologique de Wallonie (Belgique). Geologica Belgica, 15(3), 169–175.
122Berg, L. S., 1937. A classification of fish-like vertebrates. Bulletin of the Academy of Sciences of USSR, Class of Mathematics and Natural Sciences, Biological Series, 4, 1277–1280 [in English and Russian].
123Berg, L. S., 1940. Classification of fishes, both recent and fossil. Trudy Zoologicheskogo Instituta. Akademiya Nauk SSSR, Leningrad, 5, 87–517 [in Russian].
124Béthune, P. de., 1954. Carte géologique de Belgique (échelle 1/500.000). Atlas de Belgique, planche 8, Académie royale de Belgique, Bruxelles.
125Blom, H., Clack, J. A., Ahlberg, P. E. & Friedman, M., 2007. Devonian vertebrates from East Greenland: a review of faunal composition and distribution. Geodiversitas, 29(1), 119–141.
126Burrow, C. J., 2007. Early Devonian (Emsian) acanthodian faunas of the Western USA. Journal of Paleontology, 81(5), 825–840.
127Clément, G., 2002. Large Tristichopteridae (Sarcopterygii, Tetrapodomorpha) from the Late Famennian Evieux Formation of Belgium. Palaeontology, 45(3), 577–593.
128Clément, G. & Boisvert, C. A., 2006. Lohest's true and false ‘Devonian amphibians’: evidence for the rhynchodipterid lungfish Soederberghia in the Famennian of Belgium. Journal of Vertebrate Paleontology, 26(2), 276–283.
129Clément, G., Snitting, D. & Ahlberg, P. E., 2009. A new tristichopterid (Sarcopterygii, Tetrapodomorpha) from the upper Famennian Evieux Formation (Upper Devonian) of Belgium. Palaeontology, 52(4), 823–836.
130Clément, G., Ahlberg, P. E., Blieck, A., Blom, H., Clack, J. A., Poty, E., Thorez, J. & Janvier, P., 2004. Devonian tetrapod from western Europe. Nature, 427, 412–413.
131Cloutier, R., 1996. Dipnoi (Akinetia: Sarcopterygii). In Schultze, H.-P. & Cloutier, R. (eds), Devonian fishes and plants of Miguasha, Quebec, Canada. Verlag Dr. Friedrich Pfeil, München, 198–226.
132Coates, M. I. & Friedman, M., 2010. Litoptychus bryanti and characteristics of stem tetrapod neurocrania. In Elliott, D. K., Maisey, J. G., Yu, X.-B. & Miao, D. (eds), Morphology, phylogeny and paleobiogeography of fossil fishes. Verlag Dr. Friedrich Pfeil, München, 389–416.
133Cope, E. D., 1889. Synopsis of the families of Vertebrata. American Naturalist, 23, 849–877.
134Daeschler, E. B., Frumes, A. C. & Mullison, C. F., 2003. Groenlandaspidid placoderm fishes from the Late Devonian of North America. Records of the Australian Museum, 55, 45–60.
135Derycke-Khatir, C., 2005. Microrestes de vertébrés du Paléozoïque supérieur de la Manche au Rhin. Publication de la Société Géologique du Nord, 33, 1–363.
136Derycke, C. & Chancogne-Weber, C., 1995. Histological discovery on acanthodian scales from the Famennian of Belgium. In Lelièvre, H., Wenz, S., Blieck, A. & Cloutier, R. (eds), Premiers vertébrés et vertébrés inférieurs. Géobios, M. S. n°19, 31–34.
137Derycke, C. & Clément, G., 2013. First assemblage of acanthodian scales and spines from the Famennian (Upper Devonian) of Durnal (Belgium), palaeobiogeographical and palaeoenvironmental implications. Geologica Belgica, 16(1-2), 18–26.
138Fairon-Demaret, M., 1996a. Dorinnotheca streelii Fairon-Demaret, gen. et sp. nov., a new early seed plant from the upper Famennian of Belgium. Review of Palaeobotany and Palynology, 93, 217–233.
139Fairon-Demaret, M., 1996b. The plant remains from the Late Famennian of Belgium: A review. Palaeobotanist, 45, 201–208.
140Fairon-Demaret, M. & Scheckler, S. E., 1987. Typification and redescription of Moresnetia zalesskyi Stockmans, 1948, an early seed plant from the Upper Famennian of Belgium. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 57, 183–199.
141Friedman, M., 2007. The interrelationships of Devonian lungfishes (Sarcopterygii: Dipnoi) as inferred from the neurocranial evidence and new data from the genus Soederberghia Lehman, 1959. Zoological Journal of the Linnean Society, 151, 115–171.
142Gross, W., 1933. Die Fische des baltischen Devons. Palaeontographica A, 69, 1–74.
143Gross, W., 1940. Acanthodier und placodermen aus den Heterostius-Schichten Estlands und Lettlands. Annales Societatis Rebus Naturae Investigandis in Universitate Tartuensi Constitutae, 46, 12–99.
144Heintz, A., 1932. Beitrag zur Kenntnis der devonischen Fischfaunen Ost-Grønlands. Skrifter om Svalbard og Ishavet, 42, 1–27.
145Hilton, J., 1998. Review of the fossil evidence for the origin and earliest evolution of the seed-plants. Acta Botanica Sinica, 40, 981–987.
146Huxley, T. H., 1880. On the applications of the laws of evolution to the arrangement of the Vertebrata and more particularly of the Mammalia. Proceedings of the Zoological Society of London, 43, 649–662.
147Janvier, P. & Pan, J., 1982. Hyrcanaspis bliecki n. g. n.sp., a new primitive euantiarch (Antiarchi, Placodermi) from the Middle Devonian of northeastern Iran, with a discussion on antiarch phylogeny. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 164, 364–392.
148Jarvik, E., 1942. On the structure of the snout of crossopterygians and lower gnathostomes in general. Zoologiska Bidrag från Uppsala, 21, 235–675.
149Jarvik, E., 1950. Note on Middle Devonian crossopterygians from the eastern part of Gauss Halvö, East Greenland. Meddelelser om Grønland, 149(6), 1–20.
150Jarvik, E., 1972. Middle and Upper Devonian Porolepiformes from East Greenland with special reference to Glyptolepis groenlandica n. sp. and a discussion on the structure of the head in the Porolepiformes. Meddelelser om Grønland, 187(2), 1–307.
151Johanson, Z., 1997. New Remigolepis (Placodermi; Antiarchi) from Canowindra, New South Wales, Australia. Geological Magazine, 134(6), 813–846.
152Lebedev, O. A., Luksevics, E. & Zakharenko, G. V., 2010. Palaeozoogeographical connections of the Devonian vertebrate communities of the Baltica Province. Part II. Late Devonian. Palaeoworld, 19, 108–128.
153Lehman, J.-P., 1959. Les dipneustes du Dévonien supérieur du Groenland. Meddelelser om Grønland, 160(4), 1–58.
154Leriche, M., 1931. Les poissons famenniens de la Belgique - Les faciès du Famennien dans la région gallo-belge - Les relations entre les formations marines et les formations continentales du Dévonien supérieur sur la bordure méridionale du Continent Nord-Atlantique. Mémoires de la Classe des Sciences de l'Académie Royale de Belgique, in 4°, 10(5), 1–72.
155Lohest, M., 1888. Recherche sur les poissons des terrains paléozoïques de Belgique. Poissons des Psammites du Condroz, Famennien supérieur. Annales de la Société géologique de Belgique, 15, M112–M203.
156McCoy, F., 1848. On some new fossil fish of the Carboniferous period. Annals and Magazine of Natural History, 2(2), 1–10.
157Miles, R. S., 1968. The Old Red Sandstone antiarchs of Scotland: family Bothriolepididae. Monograph of the Palaeontographical Society, 122(522), 1-130.
158Miles, R. S., 1973. An actinolepid arthrodire from the Lower Devonian Peel Sound Formation, Prince of Wales Island. Palaeontographica, 143, 109–118.
159Mottequin, B., Poty, E. & Prestianni, C., 2015. Catalogue of the types and illustrated specimens recovered from the 'black marble' of Denée, a marine conservation-Lagerstätte from the Mississippian of southern Belgium. Geologica Belgica, 18(1), 1–14.
160Moy-Thomas, J. A., 1939. Palaeozoic fishes. Chemical Publing co. inc., New York, 149 pp.
161Obruchev, D. V., 1964. Osnovy paleontologii [Fundamentals of paleontology. Agnathes. Pisces]. In Orlov, J. A. (ed.), Volume 11. Nauka, Moscow, 522 pp. [in Russian].
162Olive, S., 2015. Devonian antiarch placoderms from Belgium revisited. Acta Palaeontologica Polonica, 60(3), doi: 10.4202/app.00015.2013.
163Olive, S., Prestianni, C. & Dupret, V., 2015. A new species of Groenlandaspis Heintz, 1932, (Placodermi, Arthrodira) from the Famennian (Late Devonian) of Belgium. Journal of Vertebrate Paleontology, 35(4), e935389-1 - e935389-11.
164Owen, R., 1846. Lecture on the comparative anatomy and physiology of the vertebrate animals, delivered at the Royal College of Surgeons of England in 1844 and 1846, Volume I, Fishes. Longman, Brown, Green & Longmans, London, 308 pp.
165Owen, R., 1860. Palaeontology, or a systematic summary of extinct animals and their geological relations. Black, Edinburgh, 420 pp.
166Prestianni, C., 2005. Early diversification of seeds and seed-like structures. In Steemans, P. & Javaux, E. (eds), Pre-Cambrian to Palaeozoïc Palaeopalynology and Palaeobotany. Carnets de Géologie / Notebooks on Geology, Memoir 2005/02, Abstract 06, 33-38.
167Prestianni, C. & Gerrienne, P., 2006. Lectotypification of the Famennian pre-ovule Condrusia rumex Stockmans, 1948. Review of Palaeobotany and Palynology, 142, 161–164.
168Prestianni, C. & Gerrienne, P., 2010. Early seed plant radiation: an ecological hypothesis. In: Vecoli, M., Clément, G. & Meyer-Berthaud, B. (eds). The terrestrialization process: modelling complex interactions at the biosphere-geosphere interface. Geological Society, London Special publications, 339, 71–80.
169Prestianni, C. & Gerrienne, P., 2015. Thorezia vezerensis, a new seed plant with multiovulate cupules from the Late Devonian of Belgium. Historical Biology, 27(3-4), 316-324.
170Prestianni, C., Hilton, J. & Cressler, W. L., 2013. Were all Devonian seeds cupulate? A reinvestigation of Pseudosporogonites hallei, Xenotheca bertrandii, and Aglosperma spp. International Journal of Plant Sciences, 174, 832–851.
171Prestianni, C., Caraglio, Y., Guédon, Y., Meyer-Berthaud, B. & Gerrienne, P., 2007. The seed plant Moresnetia: morphology and anatomy of the stem and fertile frond. Botany and Plant Biology 2007 Joint Congress, July 7-11, 2007, Chicago, 43-47.
172Romer, A. S., 1955. Herpetichthyes, Amphibioidei, Choanichthyes, or Sarcopterygii? Nature, 176, 126.
173Rothwell, G. W., 1986. Classifying the earliest gymnosperms. In Spicer, R. A. & Thomas, B. A. (eds), Systematic and taxonomic approaches in palaeobotany. Oxford University Press, Oxford, Systematics Association Special Volume 31, 137-161.
174Säve-Söderbergh, G., 1937. On Rhynchodipterus elginensis n. g., n. sp., representing a new group of dipnoan-like Choanata from the Upper Devonian of East Greenland and Scotland. Arkiv för Zoologi, 29(10), 1–8.
175Schultze, H.-P. & Chorn, J., 1998. Sarcopterygian and other fishes from the marine Upper Devonian of Colorado, U.S.A. Mitteilungen aus dem Museum für Naturkunde Berlin, Geowissenchaftliche Reihe, 1, 53–72.
176Serbet, R. & Rothwell, G. W., 1992. Characterising the most primitive seed ferns. I. A reconstruction of Elkinsia polymorpha. International Journal of Plant Sciences, 153, 602–621.
177Stensiö, E., 1931. Upper Devonian Vertebrates from East Greenland collected by the Danish Greenland Expeditions in 1929 and 1930. Meddelelser om Grønland, 86(1), 1-212.
178Stensiö, E., 1934. On the Placodermi of the Upper Devonian of East Greenland. I. Phyllolepida and Arthrodira. Meddelelser om Grønland, 97(1), 1–58.
179Stockmans, F., 1948. Végétaux du Dévonien supérieur de la Belgique. Mémoires du Musée Royal d'Histoire Naturelle de Belgique, 110, 1–85.
180Taverne, L., 1997. Osorioichthys marginis, "Paléonisciforme" du Famennien de Belgique, et la phylogénie des actinoptérygiens dévoniens (Pisces). Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 67, 57–78.
181Thorez, J. & Dreesen, R., 1986. A model of regressive depositional system around the Old Red Continent as exemplified by a field trip in the Upper Famennian "Psammites du Condroz" in Belgium. Annales de la Société géologique de Belgique, 109, 285–323.
182Thorez, J. & Dreesen, R., 2002. The Condroz Sandstone Group (Upper Famennian) revisited. Geologica Belgica International Meeting, Leuven, 11-15 September 2002, field trip guide-book.
183Thorez, J., Goemaere, E. & Dreesen, R., 1988. Tide- and wave-influenced depositional environments in the Psammites du Condroz (Upper Famennian), in Belgium. In De Boer, P. L., Van Gelder, A. & Nio, S. D. (eds), Tide-influenced Sedimentary Environments and Facies, D. Reidel Publishing Company, Dordrecht, 389–415.
184Thorez, J., Dreesen, R. & Streel, M., 2006. Famennian. Geologica Belgica, 9(1-2), 27–45.
185Thorez, J., Streel, M., Bouckaert, J. & Bless, M., 1977. Stratigraphie et paléogéographie de la partie orientale du Synclinorium de Dinant (Belgique) au Famennien supérieur: un modèle de bassin sédimentaire reconstitué par analyse pluridisciplinaire sédimentologique et micropaléontologique. Mededelingen rijks geologische dienst, 28, 17–32.
186Woodward, A. S., 1891. Catalogue of the fossil fishes in the British Museum of Natural History. Part II. Containing the Elasmobranchii (Acanthodii), Holocephali, Ichthyodorulites, Ostracodermi, Dipnoi, and Teleostomi (Crossopterygii), and chondrostean Actinopterygii, Volume II. British Museum of Natural History, London, 567 pp.
187Woodward, A. S., 1900. Notes on some Upper Devonian fish-remains discovered by Prof. A.G. Nathorst in East Greenland. Bihang Svenska Vetenskapsakademiens Handlinger, 26, 1–10.
188Young, G. C., 2005. An articulated phyllolepid fish (Placodermi) from the Devonian of central Australia: implications for non-marine connections with the Old Red Sandstone continent. Geological Magazine, 142(2), 173–186.
189Young, G. C. & Burrow, C. J., 2004. Diplacanthid acanthodians from the Aztec Siltstone (late Middle Devonian) of Southern Victoria Land, Antarctica. Fossils & Strata, 50, 23–43.
190Manuscript received 28.10.2014, accepted in revised form 01.02.2015, available on line 14.07.2015.